


Đau là một chức năng rất quan trọng của cơ thể con người, bao gồm sự tham gia của nociceptors và hệ thống thần kinh trung ương, hoặc CNS, để truyền tải thông điệp từ sự kích thích độc hại đến não. Nociceptors là tuyến thượng thận chịu trách nhiệm phát hiện các kích thích nguy hiểm hoặc có hại và truyền tín hiệu điện vào hệ thần kinh. Các thụ thể có mặt trong da, viscera, cơ bắp, khớp và màng não để khám phá một loạt các kích thích, có thể là cơ học, nhiệt hoặc hóa chất.
Có hai loại nociceptors:
- C-xơ sẽ là loại phổ biến nhất và chậm tiến hành và đáp ứng với kích thích. Khi các protein trong màng của thụ thể chuyển đổi sự kích thích thành những xung điện có thể được truyền qua hệ thần kinh.
- Các sợi A-delta được biết là tiến hành nhanh hơn và truyền tải thông điệp về những cơn đau dữ dội, tức thời.
Ngoài ra, có những nociceptors im lặng thường được giới hạn để kích thích nhưng có thể được "đánh thức" với kích thích cơ học cường độ cao để đáp ứng với các chất trung gian hóa học từ cơ thể. Nociceptors có thể có nhiều trạm điện áp khác nhau để truyền tải gây ra một tập hợp các hoạt động tiềm năng để bắt đầu tín hiệu điện cho hệ thần kinh. Sự kích thích và hành vi của tế bào được dựa trên các loại kênh trong nociceptor.
Điều quan trọng là phân biệt giữa nociception và đau khi xem xét cơ chế của cơn đau. Nociception là phản ứng bình thường của cơ thể để kích thích độc hại, bao gồm phản xạ dưới ngưỡng suprathreshold bảo vệ cơ thể con người khỏi bị thương. Đau chỉ được nhận thức khi superthreshold cho những người nociceptors để đạt được một hành động có thể và bắt đầu con đường đau đớn là đạt được, đó là tương đối cao. Mục đích của bài viết dưới đây là để chứng minh các cơ chế tế bào và phân tử của đau, bao gồm đau cấp tính và đau mãn tính, hoặc đau dai dẳng, như được đề cập dưới đây.
Mục lục
Cơ chế phân tử tế bào và phân tử của đau
Tóm tắt
Hệ thống thần kinh phát hiện và giải thích một loạt các kích thích nhiệt và cơ học cũng như các chất kích thích hóa học môi trường và nội sinh. Khi cường độ cao, những kích thích này tạo ra đau cấp tính và trong tình trạng chấn thương dai dẳng, cả các thành phần hệ thần kinh trung ương và ngoại vi của đường truyền đau đều thể hiện tính dẻo dai cực lớn, tăng cường tín hiệu đau và tạo ra quá mẫn. Khi dẻo tạo điều kiện cho phản xạ bảo vệ, nó có thể có lợi, nhưng khi những thay đổi vẫn tồn tại, một tình trạng đau mãn tính có thể xảy ra. Các nghiên cứu về di truyền, điện sinh lý và dược lý đang làm sáng tỏ các cơ chế phân tử làm phát hiện, mã hóa và điều chế các kích thích độc hại gây ra đau.
Giới thiệu: Đau cấp tính so với đau dai dẳng
Khả năng phát hiện các kích thích độc hại là điều cần thiết cho sự tồn tại và an sinh của sinh vật. Điều này được minh họa một cách đáng kể bằng cách kiểm tra các cá nhân bị dị tật bẩm sinh khiến họ không có khả năng phát hiện các kích thích đau đớn. Những người này không thể cảm thấy đau xuyên từ một vật sắc nhọn, sức nóng của một ngọn lửa mở, hoặc thậm chí khó chịu liên quan đến thương tích bên trong, chẳng hạn như một xương bị gãy. Kết quả là, họ không tham gia các hành vi bảo vệ thích hợp chống lại các điều kiện này, nhiều trong số đó có thể đe dọa đến tính mạng.
Thông thường, những thay đổi của con đường đau đớn dẫn đến quá mẫn cảm, vì vậy cơn đau đó sẽ vượt qua tính hữu ích của nó như là một hệ thống cảnh báo cấp tính và thay vào đó trở nên mãn tính và suy nhược. Điều này có thể được nhìn thấy, ở một mức độ nào đó, như là một phần mở rộng của quá trình chữa lành bình thường, theo đó tổn thương mô hoặc thần kinh gây ra sự hiếu động thái quá để thúc đẩy bảo vệ vùng bị thương. Ví dụ, cháy nắng tạo ra sự nhạy cảm tạm thời của khu vực bị ảnh hưởng. Kết quả là những kích thích bình thường, chẳng hạn như cảm ứng nhẹ hoặc ấm áp, được coi là đau (một hiện tượng được gọi là allodynia), hoặc kích thích đau đớn thường gây ra đau đớn cường độ lớn hơn (gọi là hyperalgesia). Ở cực đoan của nó, sự nhạy cảm không giải quyết được. Thật vậy, những người bị viêm khớp, đau dây thần kinh hậu sản (sau một cơn bệnh zona), hoặc ung thư xương, kinh nghiệm đau dữ dội và thường không ngớt mà không chỉ gây suy nhược về mặt sinh lý và tâm lý, mà còn có thể cản trở phục hồi. Đau mãn tính thậm chí có thể kéo dài lâu sau chấn thương cấp tính, có lẽ thường gặp nhất là đau lưng hoặc đau thần kinh tọa thấp hơn.
Hội chứng đau dai dẳng hoặc mãn tính có thể được bắt đầu hoặc duy trì tại loci ngoại vi và / hoặc trung tâm. Trong cả hai trường hợp, việc làm sáng tỏ các phân tử và các loại tế bào làm cho cảm giác đau bình thường (cấp tính) là chìa khóa để hiểu các cơ chế gây đau quá mẫn. Trong tổng quan hiện tại, chúng tôi nhấn mạnh sự phức tạp phân tử của các sợi thần kinh chủ yếu phát hiện các kích thích độc hại. Chúng tôi không chỉ tóm tắt việc xử lý các cơn đau cấp tính, mà còn mô tả những thay đổi trong việc xử lý đau xảy ra trong quá trình thiết lập tổn thương mô hoặc thần kinh.
Sự khác biệt sâu sắc giữa đau cấp tính và mãn tính nhấn mạnh thực tế là đau không được tạo ra bởi một hệ thống không dây, không dây, mà là kết quả từ sự tham gia của các phân tử và mạch nhựa cao, cơ sở sinh hóa phân tử và cơ sở thần kinh trong đó là trọng tâm của Các nghiên cứu hiện nay. Quan trọng hơn, thông tin mới này đã xác định được một loạt các mục tiêu điều trị tiềm năng để điều trị đau. Chúng tôi tập trung ở đây vào các tế bào thần kinh ngoại biên và thứ hai trong tủy sống; người đọc được giới thiệu đến một số đánh giá xuất sắc về cơ chế xử lý đau siêu âm, trong đó bao gồm những hiểu biết đáng chú ý mà các nghiên cứu hình ảnh đã mang đến hiện trường (Apkarian et al., 2005).
Tổng quan về giải phẫu
Sự hình thành là quá trình mà các kích thích nhiệt, cơ học hoặc hóa học cường độ cao được phát hiện bởi một quần thể nhỏ các sợi thần kinh ngoại vi, được gọi là nociceptors (Basbaum và Jessell, 2000). Các thân tế bào của cơ quan thụ cảm nằm ở hạch rễ lưng (DRG) đối với thân và hạch sinh ba đối với mặt, và có cả nhánh trục ngoại vi và trung tâm tương ứng với cơ quan đích và tủy sống của chúng. Các cơ quan thụ cảm chỉ bị kích thích khi cường độ kích thích đạt đến ngưỡng độc hại, cho thấy rằng chúng sở hữu các đặc tính lý sinh và phân tử cho phép chúng phát hiện và phản ứng có chọn lọc với các kích thích có khả năng gây hại. Có hai loại nociceptor chính. Loại thứ nhất bao gồm các liên kết được myelin hóa (A?) Có đường kính trung bình làm trung gian cho cơn đau “đầu tiên” cấp tính, khu trú tốt hoặc nhanh. Các thành phần có bao myelin này khác biệt đáng kể so với đường kính lớn hơn và dẫn truyền nhanh chóng A? các sợi đáp ứng với kích thích cơ học vô hại (tức là chạm nhẹ). Lớp nociceptor thứ hai bao gồm các sợi “C” không có bao myelin có đường kính nhỏ truyền dẫn cảm giác đau cục bộ kém, “thứ hai” hoặc đau chậm.
Các nghiên cứu điện sinh lý đã chia nhỏ hơn A? nociceptor thành hai lớp chính. Loại I (HTM: cơ quan thụ cảm cơ học ngưỡng cao) đáp ứng với cả kích thích cơ học và hóa học, nhưng có ngưỡng nhiệt tương đối cao (> 50C). Tuy nhiên, nếu kích thích nhiệt được duy trì, các hướng tâm này sẽ phản ứng ở nhiệt độ thấp hơn. Và quan trọng nhất, chúng sẽ nhạy cảm (tức là ngưỡng nhiệt hoặc cơ học sẽ giảm xuống) trong bối cảnh tổn thương mô. Loại II A? nociceptor có ngưỡng nhiệt thấp hơn nhiều, nhưng ngưỡng cơ học rất cao. Hoạt động của yếu tố hướng tâm này gần như chắc chắn làm trung gian cho phản ứng đau cấp tính “đầu tiên” đối với nhiệt độc hại. Thật vậy, khối chèn ép của các sợi thần kinh ngoại vi được myelin hóa giúp loại bỏ cơn đau đầu tiên, nhưng không phải thứ hai. Ngược lại, sợi loại I có khả năng làm trung gian cho cơn đau đầu tiên do châm chích và các kích thích cơ học cường độ cao khác gây ra.
Các sợi C không tráng myelin cũng không đồng nhất. Giống như các myelinated afferents, hầu hết các sợi C là đa thức, có nghĩa là, chúng bao gồm một dân số đó là cả nhiệt và nhạy cảm về cơ học (CMHs) (Perl, 2007). Quan tâm đặc biệt là nhiệt đáp ứng, nhưng không nhạy cảm về mặt cơ học afferents (cái gọi là nociceptors im lặng) phát triển độ nhạy cơ học chỉ trong các thiết lập của chấn thương (Schmidt et al., 1995). Những người bệnh này phản ứng nhanh hơn với kích thích hóa học (capsaicin hoặc histamine) so với CMHs, và có khả năng phát huy tác dụng khi tình trạng viêm hóa học làm thay đổi tính chất của chúng. Các tập hợp con của những người này cũng đáp ứng với nhiều loại pruritogens sản sinh ngứa. Điều đáng chú ý là không phải tất cả các sợi C đều là nociceptors. Một số phản ứng với làm mát, và một dân số đặc biệt thú vị của các nhân viên không được kết hợp với phản ứng với sự vuốt ve vô hại của da lông, nhưng không làm nóng hoặc kích thích hóa học. Những sợi sau này xuất hiện để làm trung gian cảm ứng dễ chịu (Olausson et al., 2008).
Đặc tính thần kinh và phân tử của nociceptors đã chứng minh tính không đồng nhất của chúng, đặc biệt đối với các sợi C (Snider và McMahon, 1998). Ví dụ, cái gọi là 'peptidergic' dân số của C nociceptors giải phóng các neuropeptide, chất P, và peptide liên quan đến gen calcitonin (CGRP); họ cũng biểu hiện thụ thể neurotrophin TrkA, phản ứng với yếu tố tăng trưởng thần kinh (NGF). Dân số không peptidergic của C nociceptors thể hiện các thụ thể neurotrophin c-Ret được nhắm mục tiêu bởi yếu tố thần kinh có nguồn gốc từ glial (GDNF), cũng như neurturin và artemin. Một tỷ lệ lớn dân số c-Ret dương cũng liên kết với IB4 isolectin, và biểu hiện các thụ thể kết hợp protein G của họ Mrg (Dong et al., 2001), cũng như các phân nhóm thụ thể purinergic đặc biệt, đặc biệt là P2X3. Nociceptors cũng có thể được phân biệt theo sự khác biệt giữa các kênh truyền nhiệt độ nhạy cảm (TRPV1), lạnh (TRPM8), axit chua (ASIC), và một loạt các chất kích thích hóa học (TRPA1) (Julius và Basbaum, 2001). Như đã lưu ý dưới đây, các lớp nociceptors không đồng nhất về mặt chức năng và phân tử này kết hợp với chức năng cụ thể trong việc phát hiện các phương thức đau khác nhau.
Nociceptor: Máy báo hiệu hai hướng
Người ta thường nghĩ về người nuôi cấy như mang thông tin theo một hướng, truyền các kích thích độc hại từ ngoại vi đến tủy sống. Tuy nhiên, các sợi chủ yếu có một hình thái độc đáo, được gọi là giả đơn cực, trong đó cả hai thiết bị đầu cuối trung tâm và ngoại vi phát ra từ một thân cây rìu chung. Phần lớn các protein được tổng hợp bởi DRG hoặc tế bào hạch sinh ba được phân phối cho cả hai thiết bị đầu cuối trung tâm và ngoại vi. Điều này phân biệt tế bào thần kinh chủ yếu từ nơron nguyên mẫu, nơi mà nhánh người nhận của nơron (dendrite) là khác biệt về mặt hóa học so với nhánh truyền (sợi trục). Tương đương sinh hóa của các thiết bị đầu cuối trung tâm và ngoại vi có nghĩa là nociceptor có thể gửi và nhận tin nhắn từ hai đầu. Ví dụ, cũng giống như các thiết bị đầu cuối trung tâm là locus của Ca2 + độc lập phát hành thần kinh độc lập, do đó, các thiết bị đầu cuối ngoại vi phát hành một loạt các phân tử ảnh hưởng đến môi trường mô địa phương. Viêm thần kinh, trên thực tế, đề cập đến quá trình nhờ đó phát hành ngoại vi của các neuropeptide, CGRP và chất P, gây giãn mạch và thoát mạch của protein huyết tương, tương ứng (Basbaum và Jessell, 2000). Hơn nữa, trong khi chỉ có thiết bị đầu cuối ngoại vi của nociceptor sẽ đáp ứng với kích thích môi trường (kích thích nhiệt độ, lạnh và cơ học), cả hai thiết bị đầu cuối ngoại vi và trung tâm có thể được nhắm mục tiêu bởi một loạt các phân tử nội sinh (như pH, lipid và chất dẫn truyền thần kinh) điều chỉnh độ nhạy của nó. Nó tuân theo các liệu pháp trị liệu hướng vào cả hai thiết bị đầu cuối có thể được phát triển để ảnh hưởng đến việc truyền tải thông điệp đau. Ví dụ, sự phân phối tủy sống (intrathecal) của morphine nhắm đến các thụ thể opioid được biểu hiện bởi thiết bị đầu cuối trung tâm của nociceptors, trong khi các thuốc được bôi tại chỗ (như thuốc gây tê cục bộ hoặc capsaicin) điều chỉnh cơn đau thông qua một hành động tại thiết bị ngoại vi.
Dự báo trung tâm của Nociceptor
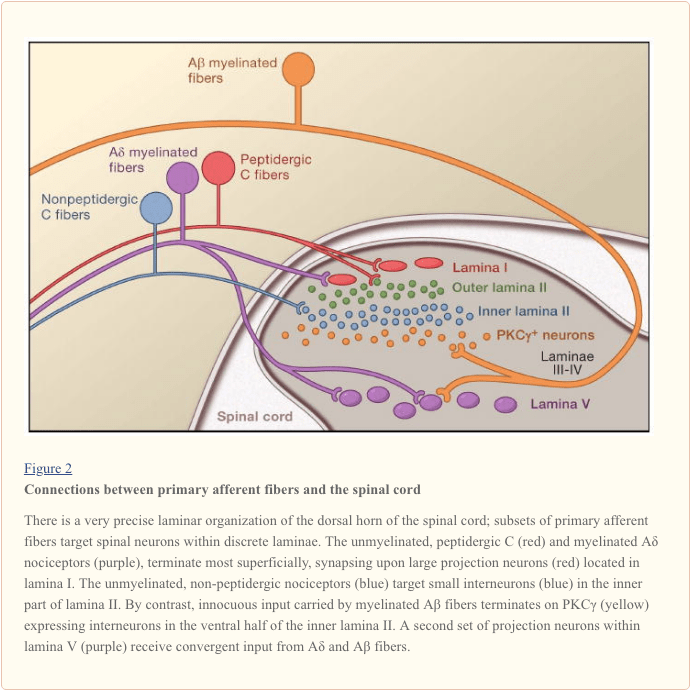
Các sợi thần kinh hướng tâm chính chiếu tới sừng lưng của tủy sống, được tổ chức thành các lớp mỏng riêng biệt về mặt giải phẫu và điện sinh lý (Basbaum và Jessell, 2000) (Hình 1). Ví dụ, A? các nociceptor chiếu vào lớp màng I cũng như lớp sừng lưng sâu hơn (lớp màng V). Ngưỡng thấp, tiến hành nhanh chóng A? hướng tâm, phản ứng với chạm nhẹ, chiếu tới các lớp sâu (III, IV và V). Ngược lại, các nociceptor C chiếu một cách hời hợt hơn tới lớp I và II.
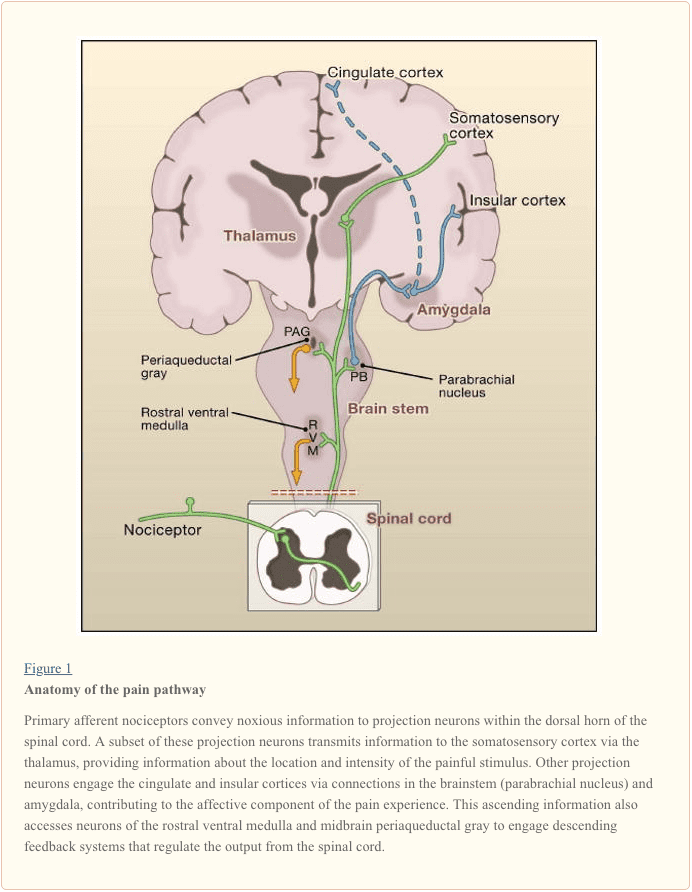
Sự phân tầng đáng chú ý này của các kiểu phụ hướng tâm bên trong sừng mặt lưng bề ngoài được làm nổi bật hơn nữa bởi các dạng hình chiếu riêng biệt của các tế bào cảm thụ C (Snider và McMahon, 1998). Ví dụ, hầu hết các sợi peptidergic C kết thúc trong lớp màng I và phần lưng nhiều nhất của lớp màng II. Ngược lại, các phần tử không thuộc hệ thống chuỗi gen, bao gồm cả tập hợp con biểu hiện Mrg, kết thúc ở vùng giữa của lamina II. Phần bụng nhất của lamina II được đặc trưng bởi sự hiện diện của các interneurons kích thích biểu hiện đồng dạng gamma của protein kinase C (PKC), có liên quan đến đau dai dẳng do chấn thương (Malmberg và cộng sự, 1997). Các nghiên cứu gần đây chỉ ra rằng PKC này? lớp được nhắm mục tiêu chủ yếu bởi các thành phần xúc cảm không cảm thụ được myelin hóa (Neumann và cộng sự, 2008). Phù hợp với các nghiên cứu giải phẫu này, các phân tích điện sinh lý chứng minh rằng các tế bào thần kinh tủy sống trong lớp màng I thường đáp ứng với kích thích độc hại (thông qua sợi A? Và C), các tế bào thần kinh trong lớp đệm III và IV chủ yếu đáp ứng với kích thích vô hại (thông qua A?), Và tế bào thần kinh trong lamina V nhận đầu vào hội tụ không độc và không độc thông qua trực tiếp (đơn âm) A? và A? đầu vào và đầu vào sợi C gián tiếp (polysynaptic). Loại thứ hai được gọi là tế bào thần kinh dải động rộng (WDR), trong đó chúng phản ứng với một loạt các cường độ kích thích. Ngoài ra còn có một đầu vào nội tạng cho các tế bào thần kinh WDR này, do đó sự hội tụ kết quả của soma và nội tạng có thể góp phần vào hiện tượng đau chuyển tiếp, theo đó đau thứ phát sau chấn thương ảnh hưởng đến mô nội tạng (ví dụ, tim bị đau thắt ngực). được đề cập đến một cấu trúc soma (ví dụ, vai).
Tăng dần các Pathways và Supraspinal Processing of Pain
Các nơron chiếu trong laminae I và V tạo thành đầu ra chính từ sừng lưng đến não (Basbaum và Jessell, 2000). Các nơron này có nguồn gốc từ nhiều con đường tăng dần, bao gồm các vùng spinothalamic và spinoreticulothalamic, mang thông điệp đau đến đồi não và não, tương ứng (Hình 2). Trước đây là đặc biệt liên quan đến các khía cạnh phân biệt cảm giác của kinh nghiệm đau (đó là, kích thích và cường độ của nó ở đâu?), Trong khi thứ hai có thể liên quan nhiều hơn đến các cơn đau cục bộ kém. Gần đây hơn, sự chú ý đã tập trung vào các dự đoán tủy sống đến vùng parabrachial của các mặt lưng dorsolateral, bởi vì đầu ra của khu vực này cung cấp một kết nối rất nhanh với amygdala, một khu vực thường được xem là xử lý thông tin liên quan đến các đặc tính của cơn đau kinh nghiệm.
Từ những bộ não và locale thalamic, thông tin đạt đến cấu trúc vỏ não. Không có vùng não đơn cần thiết cho đau (Apkarian et al., 2005). Thay vào đó, kết quả đau do kích hoạt một nhóm cấu trúc phân bố, một số kết hợp với các đặc tính phân biệt cảm giác (như vỏ não somatosensory) và những người khác với các khía cạnh cảm xúc (chẳng hạn như động mạch trước cingulate và vỏ não). Gần đây hơn, các nghiên cứu hình ảnh chứng minh sự hoạt hóa của các vùng vỏ não trước trán, cũng như các vùng không liên quan đến việc xử lý đau (như hạch bạch huyết cơ bản và tiểu não). Việc kích hoạt những vùng này có liên quan nhiều hơn đến phản ứng của cá nhân đối với kích thích hay đến nhận thức về cơn đau không rõ ràng. Cuối cùng, Hình 2 minh họa các điều khiển giảm dần mạnh mẽ có ảnh hưởng (cả tích cực và tiêu cực) sự truyền tải thông điệp đau ở mức độ của tủy sống.
Nỗi đau sâu sắc
Sợi thần kinh chủ yếu phát hiện các kích thích môi trường (của bản chất nhiệt, cơ học hoặc hóa học) và truyền tải thông tin này vào ngôn ngữ của hệ thần kinh, cụ thể là dòng điện. Đầu tiên, chúng tôi xem xét sự tiến bộ trong việc tìm hiểu cơ sở phân tử của phát hiện tín hiệu, và làm theo điều này với một tổng quan ngắn gọn về các nghiên cứu di truyền gần đây làm nổi bật sự đóng góp của các kênh điện áp đến truyền đau (Hình 3).
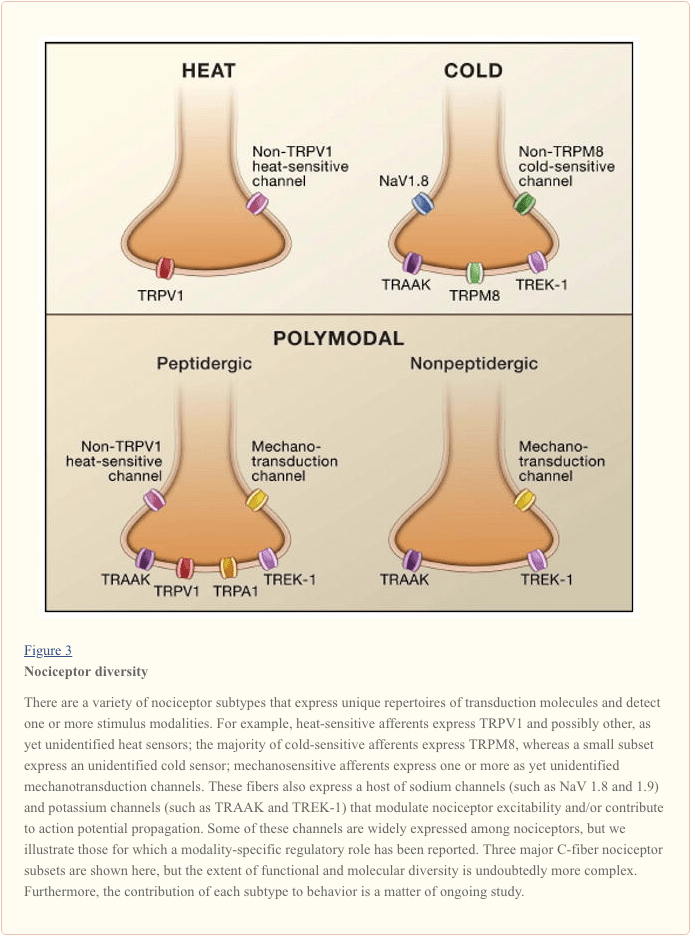
Kích hoạt Nociceptor: Nhiệt
Các nghiên cứu tâm sinh lý con người đã chỉ ra rằng có một ranh giới rõ ràng và có thể tái tạo được giữa nhận thức về hơi ấm vô hại và nhiệt độc hại, cho phép chúng ta nhận biết và tránh nhiệt độ có khả năng gây tổn thương mô. Ngưỡng đau này, thường nằm ở khoảng 43 ° C, tương đương với độ nhạy nhiệt của C và Loại II A? nociceptors được mô tả trước đó. Thật vậy, các tế bào thần kinh được nuôi cấy từ các hạch rễ lưng phân ly cho thấy độ nhạy nhiệt tương tự. Đa số hiển thị ngưỡng 43 ° C, với một nhóm nhỏ hơn được kích hoạt bởi nhiệt độ cao hơn (ngưỡng> 50 ° C) (Cesare và McNaughton, 1996; Kirschstein và cộng sự, 1997; Leffler và cộng sự, 2007; Nagy và Rang , 1999). Những hiểu biết về phân tử về quá trình cảm ứng nhiệt đến từ việc nhân bản và mô tả chức năng của thụ thể đối với capsaicin, thành phần cay chính trong ớt 'cay'. Capsaicin và các hợp chất vanilloid liên quan tạo ra cơn đau rát bằng cách khử cực các tập hợp con cụ thể của C và A? nociceptor thông qua việc kích hoạt thụ thể capsaicin (hoặc vanilloid), TRPV1, một trong khoảng 30 thành viên của họ kênh ion tiềm năng thụ thể thoáng qua (TRP) lớn hơn (Caterina et al., 1997). Kênh TRPV1 nhân bản cũng được điều chỉnh khi nhiệt độ môi trường tăng lên, với ngưỡng kích hoạt nhiệt (? 43 ° C).
Một số dòng chứng cứ ủng hộ giả thuyết rằng TRPV1 là bộ chuyển đổi nội sinh của nhiệt độc hại. Đầu tiên, TRPV1 được thể hiện ở phần lớn các nociceptors nhạy nhiệt (Caterina et al., 1997). Thứ hai, các dòng capsaicin và nhiệt kích thích tương tự nhau, nếu không giống nhau, liên quan đến các đặc tính dược lý và sinh lý của chúng, cũng giống như các dòng TRPV1 được biểu hiện dị biệt. Thứ ba, và như được mô tả chi tiết hơn dưới đây, phản ứng TRPV1 gợi lên được tăng cường rõ rệt bởi các tác nhân giảm đau hoặc pro-viêm (như proton ngoại bào, neurotrophin, hoặc bradykinin), tất cả đều tạo ra quá mẫn cảm với nhiệt in vivo (Tominaga et al., 1998)). Thứ tư, phân tích những con chuột thiếu kênh ion này không chỉ tiết lộ một sự mất mát hoàn toàn về độ nhạy capsaicin, nhưng những con vật này cũng thể hiện sự suy giảm đáng kể khả năng phát hiện và phản ứng với nhiệt độc hại (Caterina và cộng sự, 2000; Davis et al., 2000 ). Những nghiên cứu này cũng đã chứng minh vai trò thiết yếu cho kênh này trong quá trình nhờ đó tổn thương mô và viêm dẫn đến quá mẫn cảm nhiệt, phản ánh khả năng của TRPV1 là chất tích hợp phân tử của kích thích nhiệt và hóa học (Caterina et al., 2000; Davis et al ., 2000).
Tuy nhiên, sự đóng góp của TRPV1 vào cảm giác nhiệt cấp tính đã được thử thách bởi dữ liệu thu được từ một chế phẩm ex vivo trong đó ghi nhận được từ soma của các tế bào thần kinh DRG với các sợi trung tâm và ngoại vi nguyên vẹn. Trong một nghiên cứu, không có sự khác biệt nào được quan sát thấy trong phản ứng nhiệt từ loại hoang dã và động vật thiếu TRPV1 (Woodbury et al., 2004), nhưng một phân tích gần đây hơn từ nhóm này cho thấy những con chuột thiếu TRPV1 thực sự thiếu nhóm thuần tập của tế bào thần kinh được kích hoạt mạnh mẽ bởi nhiệt độc hại (Lawson và cộng sự, 2008). Kết hợp với các kết quả được mô tả ở trên, chúng tôi kết luận rằng TRPV1 không thể nghi ngờ góp phần vào cảm giác nhiệt cấp tính, nhưng đồng ý rằng TRPV1 không chỉ chịu trách nhiệm về sự truyền nhiệt.
Về mặt này, trong khi các con chuột thiếu TRPV1 thiếu một thành phần nhạy cảm với nhiệt độ hành vi, việc sử dụng capsaicin liều cao để loại bỏ các thiết bị đầu cuối trung tâm của TRPV1 thể hiện các xơ chủ yếu gây ra sự mất mát nghiêm trọng hơn. (Cavanaugh và cộng sự, 2009). Đối với đột biến TRPV1, cũng có sự mất mát của tăng cường nhiệt gây tổn thương mô. Kết hợp các kết quả này cho thấy cả thành phần độc lập TRPV1 và TRPV1 độc lập với độ nhạy nhiệt độc hại đều được trung gian thông qua các nociceptors TRPV1.
Điều gì tạo nên thành phần độc lập với TRPV1 của cảm giác nhiệt? Một số loại kênh TRPV phụ khác, bao gồm TRPV2, 3 và 4, đã nổi lên như các đầu dò nhiệt ứng cử viên có khả năng bao gồm việc phát hiện các cường độ kích thích bên cạnh TRPV1, bao gồm cả rất nóng (> 50 ° C) và ấm (giữa 30 ° Cs) nhiệt độ (Lumpkin và Caterina, 2007). Các kênh TRPV2 được biểu hiện khác nhau hiển thị ngưỡng kích hoạt nhiệt độ là 52 ° C, trong khi TRPV3 và TRPV4 được kích hoạt trong khoảng 25 - 35 ° C. TRPV2 được thể hiện trong một quần thể con của A? các tế bào thần kinh phản ứng với nhiệt độc hại ngưỡng cao và các đặc tính lý sinh của nó giống với các đặc tính của dòng điện kích thích nhiệt ngưỡng cao nguyên bản (Leffler và cộng sự, 2007; Rau và cộng sự, 2007). Chưa có báo cáo nào được công bố mô tả các thử nghiệm sinh lý hoặc hành vi của chuột loại trực tiếp TRPV2. Mặt khác, những con chuột thiếu TRPV3- và TRPV4 hiển thị sở thích nhiệt bị thay đổi khi được đặt trên bề mặt có nhiệt độ phân cấp, cho thấy rằng các kênh này góp phần theo một cách nào đó vào việc phát hiện nhiệt độ in vivo (Guler và cộng sự, 2002). Điều thú vị là cả TRPV3 và TRPV4 đều cho thấy biểu hiện ở tế bào sừng và tế bào biểu mô lớn hơn đáng kể so với tế bào thần kinh cảm giác, nâng cao khả năng phát hiện các kích thích nhiệt vô hại liên quan đến tác động qua lại chức năng giữa da và các sợi hướng tâm chính bên dưới (Chung và cộng sự, 2003; Peier và cộng sự, 2002b).
Kích hoạt Nociceptor: Lạnh
Đối với capsaicin và TRPV1, các chất làm mát tự nhiên, chẳng hạn như tinh dầu bạc hà và eucalyptol, đã được sử dụng làm chất thăm dò dược lý để xác định và mô tả các sợi và tế bào nhạy cảm với lạnh (Hensel và Zotterman, 1951; Reid và Flonta, 2001) và các phân tử làm cơ sở hành vi của họ. Thật vậy, hầu hết các tế bào thần kinh nhạy cảm với lạnh đều phản ứng với tinh dầu bạc hà và hiển thị ngưỡng kích hoạt nhiệt là 25 ° C. TRPM8 là một kênh nhạy cảm với tinh dầu bạc hà và tinh dầu bạc hà có đặc điểm sinh lý phù hợp với đặc điểm sinh lý của dòng điện lạnh bản địa và những con chuột thiếu TRPM8 cho thấy sự mất mát rất đáng kể của tinh dầu bạc hà và các phản ứng kích thích lạnh ở cấp độ tế bào hoặc sợi thần kinh. Tương tự như vậy, những con vật này thể hiện sự thiếu hụt nghiêm trọng trong các phản ứng hành vi do lạnh (Bautista và cộng sự, 2007; Colburn và cộng sự, 2007; Dhaka và cộng sự, 2007) trong một phạm vi nhiệt độ rộng từ 30 đến 10 ° C. Như trong trường hợp của TRPV1 và anh ta, những con chuột thiếu TRPM8 không hoàn toàn vô cảm với lạnh. Ví dụ, vẫn còn một nhóm nhỏ (? 4%) gồm các tế bào thần kinh nhạy cảm với lạnh, nhạy cảm với tinh dầu bạc hà có ngưỡng kích hoạt thấp, khoảng 12 ° C. Những điều này có thể giải thích cho độ nhạy cảm với lạnh còn sót lại được thấy trong các thử nghiệm hành vi, trong đó động vật thiếu TRPM8 vẫn có thể tránh các bề mặt cực lạnh dưới 10 ° C. Điều quan trọng là những con chuột thiếu TRPM8 cho thấy sự nhạy cảm bình thường với nhiệt độc hại. Thật vậy, TRPV1 và TRPM8 được biểu hiện trong các quần thể tế bào thần kinh phần lớn không chồng chéo, phù hợp với quan điểm cho rằng các cơ chế phát hiện nóng và lạnh được tổ chức thành các 'đường được dán nhãn' riêng biệt về mặt giải phẫu và chức năng.
Dựa trên các hệ thống biểu hiện dị thể, TRPA1 cũng đã được đề xuất để phát hiện cảm lạnh, đặc biệt trong phạm vi độc hại (<15 ° C). Hơn nữa TRPA1 được kích hoạt bởi các hợp chất làm mát icilin và tinh dầu bạc hà (Bandell và cộng sự, 2004; Karashima và cộng sự, 2007; Story và cộng sự, 2003), mặc dù ở nồng độ tương đối cao so với tác dụng của chúng ở TRPM8. Tuy nhiên, vẫn tiếp tục có sự bất đồng về việc liệu TRPA1 bản địa hay tái tổ hợp là bản chất nhạy cảm với lạnh (Bandell và cộng sự, 2004; Jordt và cộng sự, 2004; Karashima và cộng sự, 2009; Nagata và cộng sự, 2005; Zurborg và cộng sự ., 2007). Cuộc tranh cãi này vẫn chưa được giải quyết bằng việc phân tích hai dòng chuột độc lập thiếu TRPA1. Ở cấp độ tế bào, một nghiên cứu đã chỉ ra các phản ứng bình thường do lạnh ở các tế bào thần kinh thiếu TRPA1 sau khi nhiệt độ giảm từ 30 ° C xuống 22 ° C trong 4 giây (Bautista và cộng sự, 2006); một nghiên cứu gần đây hơn đã chỉ ra sự giảm các tế bào thần kinh nhạy cảm với lạnh từ 26% (WT) xuống 10% (TRPA1 - / -), khi được thử nghiệm sau 200 giây giảm nhiệt độ, từ 30 ° C xuống 10 ° C (Karashima et al. ., 2009). Trong các nghiên cứu về hành vi, những con chuột thiếu TRPA1 thể hiện các phản ứng tương tự như các đồng loại hoang dã trong các thử nghiệm làm mát bay hơi bằng đĩa lạnh và axeton (Bautista et al., 2006). Một nghiên cứu thứ hai sử dụng các xét nghiệm tương tự cho thấy rằng động vật loại trực tiếp TRPA1 cái, chứ không phải đực, thể hiện sự nhạy cảm với lạnh kém hơn so với các động vật cùng lứa hoang dã (Kwan et al., 2006). Karashima và cộng sự không tìm thấy sự khác biệt nào về độ trễ run rẩy hoặc rút chân ở chuột đực hoặc chuột cái thiếu TRPA1 trong thử nghiệm trên tấm lạnh, nhưng quan sát thấy rằng việc tiếp xúc lâu với bề mặt lạnh khiến động vật hoang dã nhảy lên chứ không phải động vật thiếu TRPA1 (Karashima et al., 2009). Có thể hình dung, kiểu hình thứ hai phản ánh sự đóng góp của TRPA1 đối với sự nhạy cảm với lạnh trong trường hợp tổn thương mô, nhưng không phải đối với cơn đau lạnh cấp tính. Phù hợp với giả thuyết thứ hai, các bản ghi âm sợi thần kinh đơn không cho thấy sự giảm độ nhạy cảm cấp tính ở chuột thiếu TRPA1 (Cavanaugh và cộng sự, 2009; Kwan và cộng sự, 2009). Cuối cùng, đáng chú ý là những con chuột được điều trị bằng capsaicin thiếu các đầu cuối trung tâm của các sợi biểu hiện TRPV1 cho thấy các phản ứng hành vi nguyên vẹn đối với các kích thích lạnh và độc hại (Cavanaugh và cộng sự, 2009). Vì TRPA1 được biểu hiện trong một tập hợp con các tế bào thần kinh dương tính TRPV1, nên TRPA1 không cần thiết cho độ nhạy cảm lạnh cấp tính bình thường. Các nghiên cứu trong tương lai sử dụng chuột thiếu cả TRPM8 và TRPA1 sẽ giúp giải quyết những vấn đề này và xác định các phân tử và loại tế bào làm cơ sở cho thành phần độc lập TRPM8 còn sót lại của độ nhạy cảm lạnh.
Các phân tử bổ sung, bao gồm các kênh natri điện áp (được thảo luận bên dưới), các kênh kali điện áp và các kênh kali KCNK hai lỗ, phối hợp với TRPM8 để tinh chỉnh các ngưỡng lạnh hoặc truyền các tiềm năng hành động lạnh (Viana et al. , 2002; Zimmermann et al., 2007; Noel và cộng sự, 2009). Ví dụ, các chất ức chế Kv1 cụ thể làm tăng ngưỡng nhiệt độ của các nơron nhạy cảm với lạnh và tiêm các chất ức chế này vào hindpaw của động vật gặm nhấm làm giảm phản ứng với cảm lạnh, nhưng không làm nóng hoặc kích thích cơ học (Madrid và cộng sự, 2009). Hai thành viên của họ kênh KCNK, KCNK2 (TREK-1) và KCNK4 (TRAAK) được thể hiện trong một tập con của các nhà nghiên cứu chất xơ C (Noel et al., 2009) và có thể được điều chế bởi nhiều kích thích sinh lý và dược lý, bao gồm cả áp lực và nhiệt độ. Hơn nữa, những con chuột thiếu các kênh này cho thấy sự bất thường về độ nhạy với áp suất, nhiệt và lạnh (Noel et al., 2009). Mặc dù những phát hiện này gợi ý rằng các kênh TREK-1 và TRAAK điều chỉnh kích thích nociceptor, vẫn chưa rõ độ nhạy nội tại của chúng đối với kích thích vật lý liên quan đến sự đóng góp trong cơ thể của chúng đối với sự truyền nhiệt hoặc cơ học.
Kích hoạt Nociceptor: Cơ
Hệ thống somatosensory phát hiện các kích thích cơ học đa dạng về số lượng và chất lượng, từ cọ nhẹ trên da đến căng phồng của thành bàng quang. Một loạt các phân nhóm thần kinh cảm ứng cơ học được chuyên biệt để phát hiện chuỗi kích thích cơ học đa dạng này và có thể được phân loại theo độ nhạy ngưỡng. Cơ quan thụ cảm ở ngưỡng cao bao gồm sợi C và sợi A thích nghi chậm? Các sợi cơ học (AM), cả hai đều kết thúc như các đầu dây thần kinh tự do trên da. Cơ chế nhận cảm ngưỡng thấp bao gồm A? Sợi lông D chấm dứt trên lông tơ trên da và phát hiện ra khi chạm nhẹ. Cuối cùng, A? các sợi bên trong tế bào Merkel, tiểu thể Pacinian và nang lông phát hiện kết cấu, độ rung và áp suất ánh sáng.
Như trong trường hợp kích thích nhiệt, độ nhạy cơ học đã được khảo sát ở một số cấp độ, bao gồm các nơ-ron cảm giác phân ly trong nuôi cấy, ghi hình sợi ex-vivo, cũng như ghi âm từ trung tâm (tức là nơ-ron sừng lưng) và các phép đo đầu ra hành vi. Các bản ghi thần kinh da ex-vivo có nhiều thông tin nhất trong việc kết hợp các đặc tính kích thích (chẳng hạn như cường độ, tần số, tốc độ và sự thích ứng) với các loại sợi phụ cụ thể. Ví dụ, A? sợi cơ bản liên quan đến độ nhạy cảm với ánh sáng, trong khi C và A? sợi chủ yếu đáp ứng với các xúc phạm cơ học độc hại. Ở cấp độ hành vi, độ nhạy cơ học thường được đánh giá bằng hai kỹ thuật. Phổ biến nhất là đo phản ứng phản xạ với lực liên tục tác dụng lên chân sau của loài gặm nhấm bằng các sợi đã được hiệu chỉnh (lông Von Frey). Loại thứ hai tác dụng áp lực tăng dần lên chân hoặc đuôi thông qua một hệ thống kẹp. Trong cả hai trường hợp, thông tin về ngưỡng cơ học được thu thập trong các tình huống bình thường (cấp tính) hoặc chấn thương (quá mẫn). Một trong những thách thức trong lĩnh vực này là phát triển các thử nghiệm hành vi bổ sung để đo lường các khía cạnh khác nhau của quá trình cảm ứng cơ học, chẳng hạn như phân biệt kết cấu và độ rung, điều này sẽ tạo điều kiện thuận lợi cho việc nghiên cứu cả chạm độc hại và không độc hại (Wetzel và cộng sự, 2007).
Ở cấp độ tế bào, áp lực có thể được áp dụng cho các tế bào của các tế bào thần kinh somatosensory (hoặc đến nơ ron) bằng cách sử dụng đầu dò thủy tinh, thay đổi cường độ thẩm thấu, hoặc kéo dài thông qua sự căng thẳng của bề mặt cấy đàn hồi, mặc dù không rõ bắt chước áp lực sinh lý (Bhattacharya et al., 2008; Cho et al., 2006; Cho et al., 2002; Drew et al., 2002; Hu và Lewin, 2006; Lin và cộng sự, 2009; Takahashi và Gotoh, 2000 ). Phản ứng có thể được đánh giá bằng cách sử dụng phương pháp hình ảnh tế bào sinh lý hoặc trực tiếp. Sự đồng thuận từ các nghiên cứu như vậy là áp suất đó mở ra một kênh cation cơ học để thu hút sự khử cực nhanh chóng. Tuy nhiên, một số ít các đầu dò dược lý cụ thể và các marker phân tử để mô tả các phản ứng này hoặc ghi nhãn các phân nhóm thần kinh có liên quan đã cản trở các nỗ lực phù hợp với các hoạt động tế bào với các phân lớp sợi thần kinh giải phẫu hoặc được xác định chức năng. Những hạn chế này cũng đã cản trở việc phân tích phân tử của mechansosensation và xác định các phân tử cấu thành máy móc cơ khí truyền tải. Tuy nhiên, một số ứng cử viên đã nổi lên, phần lớn dựa trên các nghiên cứu về cơ chế hóa trong mô hình sinh vật di truyền. Các orthologues của các protein này đã được kiểm tra bằng cách sử dụng các phương pháp nhắm mục tiêu gen ở chuột, trong đó các kỹ thuật được đề cập ở trên có thể được sử dụng để đánh giá thâm hụt cơ học ở tất cả các cấp. Dưới đây chúng tôi tóm tắt ngắn gọn một số ứng cử viên được tiết lộ trong các nghiên cứu này.
Ứng viên cơ chế ứng cử viên: Kênh DEG / ENaC
Các nghiên cứu ở tuyến trùng Caenorhabditis elegans (C. elegans) đã xác định mec-4 và mec-10, thành viên của họ degenerin / epithelial Na + channel (DEG / ENaC), như cơ chế vận chuyển trong cơ thể thần kinh cảm ứng (Chalfie, 2009). Dựa trên những nghiên cứu này, các orthologues của động vật có vú ASIC 1, 2 và 3 đã được đề xuất như các kênh truyền dẫn cơ học. ASIC là các kênh ion nhạy cảm với axit phục vụ như các thụ thể cho các proton ngoại bào (nhiễm toan mô) được tạo ra trong quá trình thiếu máu cục bộ (xem bên dưới). Mặc dù các kênh này được thể hiện bởi cả các tế bào thần kinh cơ học ngưỡng thấp và ngưỡng cao, các nghiên cứu di truyền không đồng nhất hỗ trợ một vai trò thiết yếu trong việc vận chuyển cơ học. Chuột thiếu các kênh ASIC1 chức năng hiển thị các phản ứng hành vi bình thường đối với cảm ứng da, và ít hoặc không có thay đổi về độ nhạy cơ học khi được đánh giá bằng cách ghi một sợi quang (Page et al., 2004; Price et al., 2000). Tương tự như vậy, các sợi thần kinh ngoại biên từ các con chuột thiếu ASIC2 chỉ làm giảm nhẹ khả năng hoạt động của cơ thể, trong khi các sợi thiếu ASIC3 cho thấy sự tăng nhẹ (không có sự thay đổi về ngưỡng cơ học hoặc độ nhạy cơ học hành vi cơ bản). Giá et al., 2001; Roza và cộng sự, 2004). Phân tích những con chuột thiếu cho cả ASIC2 và ASIC3 cũng không hỗ trợ vai trò của các kênh này trong quá trình vận chuyển cơ học bằng da (Drew và cộng sự, 2004). Vì vậy, mặc dù các kênh này dường như đóng một vai trò trong đau cơ xương khớp và thiếu máu cục bộ (xem bên dưới), sự đóng góp của họ đối với việc cơ khí hóa vẫn chưa được giải quyết.
Các nghiên cứu di truyền cho thấy rằng các kênh mec-4 / mec-10 của C. elegans tồn tại trong một phức hợp với protein giống stomatin MEC-2 (Chalfie, 2009). Chuột thiếu trực giao MEC-2, SLP3, hiển thị mất độ nhạy cơ học ở ngưỡng A thấp? và A? sợi, nhưng không ở dạng sợi C (Wetzel và cộng sự, 2007). Những con chuột này biểu hiện sự nhạy bén của xúc giác bị thay đổi, nhưng hiển thị phản ứng bình thường với áp lực độc hại, cho thấy rằng SLP3 góp phần phát hiện các kích thích cơ học vô hại, nhưng không độc hại. Liệu SLP3 có hoạt động trong một phức hợp truyền cơ học hay tương tác với ASIC trong tế bào thần kinh cảm giác của động vật có vú hay không vẫn chưa được biết.
Cơ chế vận động viên ứng cử viên: Kênh TRP
Như đã nói ở trên, khi được biểu hiện một cách dị dạng, TRPV2 không chỉ phản ứng với nhiệt độc hại mà còn phản ứng với độ thẩm thấu. Ngoài ra, các kênh TRPV2 bản địa trong các tế bào cơ trơn mạch máu được kích hoạt bằng cách hút trực tiếp và kích thích thẩm thấu (Muraki và cộng sự, 2003). Vai trò của TRPV2 đối với việc vận chuyển cơ học somatosensory trong cơ thể vẫn chưa được thử nghiệm.
TRPV2 được thể hiện rõ ràng ở đường kính vừa và lớn, A? sợi đáp ứng với cả kích thích cơ học và nhiệt (Caterina et al., 1999; Muraki et al., 2003). TRPV4 biểu hiện khiêm tốn ở hạch cảm giác, nhưng biểu hiện nhiều hơn ở thận và các tế bào niệu quản nhạy cảm với căng của bàng quang (Gevaert và cộng sự, 2007; Mochizuki và cộng sự, 2009). Khi biểu hiện khác nhau, cả TRPV2 và TRPV4 đều được chứng minh là phản ứng với những thay đổi của áp suất thẩm thấu (Guler và cộng sự, 2002; Liedtke và cộng sự, 2000; Mochizuki và cộng sự, 2009; Strotmann và cộng sự, 2000). Phân tích động vật thiếu TRPV4 cho thấy một vai trò trong quá trình thẩm thấu vì động vật loại trực tiếp có biểu hiện khiếm khuyết về huyết áp, cân bằng nước và độ rỗng bàng quang (Gevaert et al., 2007; Liedtke và Friedman, 2003). Những con vật này biểu hiện sự tăng trương lực cơ ở da cấp tính bình thường, nhưng có biểu hiện thiếu hụt trong các mô hình tăng trương lực cơ và nhiệt (Alessandri-Haber và cộng sự, 2006; Chen và cộng sự, 2007; Grant và cộng sự, 2007; Suzuki và cộng sự, 2003). Do đó, TRPV4 không có khả năng đóng vai trò là chất dẫn truyền cơ học chính trong các tế bào thần kinh cảm giác, nhưng có thể góp phần gây ra quá mẫn cảm với cơn đau do chấn thương.
TRPA1 cũng đã được đề xuất để phục vụ như một máy dò các kích thích cơ học. TRPA1 của động vật có vú biểu hiện dị thường được kích hoạt bởi các nhà tạo màng (Hill và Schaefer, 2007) và trực hệ giun nhạy cảm với áp suất cơ học được áp dụng thông qua một pipet hút (Kindt và cộng sự, 2007). Tuy nhiên, những con chuột thiếu TRPA1 chỉ thể hiện những khiếm khuyết yếu trong hành vi cảm ứng cơ học và kết quả không nhất quán. Hai nghiên cứu báo cáo không có thay đổi về ngưỡng cơ học ở động vật thiếu TRPA1 (Bautista và cộng sự, 2006; Petrus và cộng sự, 2007), trong khi nghiên cứu thứ ba báo cáo thiếu hụt (Kwan và cộng sự, 2006). Một nghiên cứu gần đây hơn cho thấy rằng C và A? Các sợi nhạy cảm cơ học ở động vật loại trực tiếp TRPA1 đã thay đổi phản ứng với kích thích cơ học (một số tăng lên và một số khác giảm) (Kwan và cộng sự, 2009). Các tác động sinh lý khác biệt này có biểu hiện ở mức độ hành vi hay không và như thế nào vẫn chưa rõ ràng. Tổng hợp lại, TRPA1 dường như không hoạt động như một máy dò chính của các kích thích cơ học cấp tính, nhưng có lẽ điều chỉnh khả năng kích thích của các hướng cảm cơ học.
Ứng viên cơ chế ứng viên: Kênh KCNK
Ngoài vai trò đầu dò cơ học tiềm năng của KCNK2 và 4 (xem ở trên), KCNK18 đã được thảo luận về khả năng đóng góp của nó trong việc cảm biến cơ học. Do đó, KCNK18 được nhắm mục tiêu bởi hydroxy-a-sanshool, thành phần cay nồng trong hạt tiêu Tứ Xuyên tạo ra cảm giác ngứa ran và tê liệt, gợi ý về sự tương tác với các tế bào thần kinh nhạy cảm (Bautista et al., 2008; Bryant and Mezine, 1999; Sugai et al., 2005). KCNK18 được thể hiện trong một tập hợp con của các sợi peptidergic C giả định và các cơ quan thụ cảm cơ học ngưỡng thấp (A?), Nơi nó đóng vai trò là cơ quan điều chỉnh chính về thời gian tiềm năng hoạt động và khả năng kích thích (Bautista và cộng sự, 2008; Dobler và cộng sự, 2007). Hơn nữa, sanshool khử cực các tế bào thần kinh cảm giác đường kính lớn nhạy cảm cơ học và cơ học, cũng như một tập hợp con của các tế bào thụ cảm (Bautista và cộng sự, 2008; Bhattacharya và cộng sự, 2008). Mặc dù không biết KCNK18 có nhạy cảm trực tiếp với kích thích cơ học hay không, nhưng nó có thể là một bộ điều chỉnh quan trọng đối với sự kích thích của các tế bào thần kinh liên quan đến cảm giác chạm vô hại hoặc độc hại.
Tóm lại, cơ sở phân tử của quá trình vận chuyển cơ học động vật có vú không được làm rõ. Quá mẫn cơ học để đáp ứng với mô hoặc tổn thương thần kinh là một vấn đề lâm sàng chính và do đó làm sáng tỏ cơ sở sinh học của cảm ứng trong điều kiện bình thường và sinh lý bệnh vẫn là một trong những thách thức chính trong nghiên cứu đau đớn và đau đớn.
Kích hoạt Nociceptor: Hóa chất
Chemo-nociception là quá trình mà các tế bào thần kinh chủ yếu phát hiện các chất kích thích môi trường và các yếu tố nội sinh do stress sinh lý gây ra. Trong bối cảnh đau cấp tính, các cơ chế hóa học-nociceptive gây ra phản ứng nghịch với một loạt các chất kích thích môi trường. Ở đây, một lần nữa, các kênh TRP có vai trò nổi bật, có lẽ không đáng ngạc nhiên khi chúng hoạt động như các thụ thể cho các chất kích thích có nguồn gốc từ thực vật, bao gồm capsaicin (TRPV1), menthol (TRPM8), cũng như các thành phần hăng trong cây mù tạt và tỏi, isothiocyanates và thiosulfinates (TRPA1) (Bandell và cộng sự, 2004; Caterina và cộng sự, 1997; Jordt và cộng sự, 2004; McKemy và cộng sự, 2002; Peier và cộng sự, 2002a).
Đối với các chất gây kích ứng môi trường, TRPA1 đã nổi lên như một thành viên đặc biệt thú vị của nhóm này. Điều này là do TRPA1 phản ứng với các hợp chất đa dạng về cấu trúc nhưng thống nhất về khả năng tạo cộng hóa trị với các nhóm thiol. Ví dụ, allyl isothiocyanate (từ wasabi) hoặc allicin (từ tỏi) là các electrophin thấm qua màng kích hoạt TRPA1 bằng cách biến đổi cộng hóa trị các gốc cysteine trong vùng tế bào chất đầu cuối amin của kênh (Hinman và cộng sự, 2006; Macpherson và cộng sự, 2007). Điều này thúc đẩy kiểm soát kênh như thế nào hiện vẫn chưa được biết. Tuy nhiên, chỉ cần thiết lập tầm quan trọng của phản ứng thiol trong quá trình này đã cho thấy TRPA1 là mục tiêu sinh lý chính cho một nhóm chất độc môi trường đa dạng và đa dạng về mặt hóa học. Một ví dụ đáng chú ý là acrolein (2-propenal), một chất phản ứng mạnh ?,? - aldehyde không bão hòa có trong hơi cay, khói xe, hoặc khói từ việc đốt thực vật (tức là cháy rừng và thuốc lá). Acrolein và các chất kích ứng dễ bay hơi khác (như hypochlorite, hydrogen peroxide, formalin và isocyanates) kích hoạt các tế bào thần kinh cảm giác kích hoạt mắt và đường hô hấp, gây ra đau và viêm (Bautista và cộng sự, 2006; Bessac và Jordt, 2008; Caceres và cộng sự. , 2009). Hành động này có thể gây ra những hậu quả đặc biệt nghiêm trọng đối với những người bị hen suyễn, ho mãn tính hoặc các bệnh rối loạn phổi khác. Những con chuột thiếu TRPA1 cho thấy độ nhạy cảm giảm đáng kể với các tác nhân như vậy, nhấn mạnh bản chất quan trọng của kênh này như một máy dò cảm giác của các chất kích ứng môi trường phản ứng (Caceres và cộng sự, 2009). Ngoài các chất độc từ môi trường này, TRPA1 được nhắm mục tiêu bởi một số thuốc gây mê nói chung (như isofluorane) hoặc các sản phẩm phụ chuyển hóa của các tác nhân hóa trị liệu (như cyclophosphamide), có khả năng gây ra một số tác dụng phụ bất lợi của những loại thuốc này, bao gồm đau cấp tính và viêm thần kinh mạnh. (Bautista và cộng sự, 2006; Matta và cộng sự, 2008).
Cuối cùng, các chất kích thích hóa học và các tác nhân giảm đau khác cũng được sản xuất nội sinh để đáp ứng với tổn thương mô hoặc stress sinh lý, bao gồm cả stress oxy hóa. Các yếu tố như vậy có thể hoạt động độc lập, hoặc kết hợp, để kích thích các nociceptors với các kích thích nhiệt và / hoặc cơ học, do đó làm giảm ngưỡng đau. Kết quả của hành động này là để tăng cường bảo vệ và bảo vệ phản xạ trong hậu quả của chấn thương. Do đó, hóa học-nociception đại diện cho một giao diện quan trọng giữa đau cấp tính và dai dẳng, đặc biệt là trong bối cảnh tổn thương mô ngoại vi và viêm, như được thảo luận chi tiết hơn dưới đây.
Đau cấp tính: Tiến hành tín hiệu đau
Một khi tín hiệu nhiệt và cơ học được đầu dò bởi thiết bị đầu cuối chính, tiềm năng receptor sẽ kích hoạt một loạt các kênh ion điện áp. Các kênh natri và kali có điện áp rất quan trọng đối với việc tạo ra các tiềm năng hành động truyền tín hiệu nociceptor tới các khớp thần kinh ở sừng lưng. Các kênh canxi có điện áp đóng một vai trò quan trọng trong việc giải phóng chất dẫn truyền thần kinh từ các đầu cuối nociceptor trung tâm hoặc ngoại vi để tạo ra đau hoặc viêm neurogenic, tương ứng. Chúng tôi hạn chế cuộc thảo luận của chúng tôi với các thành viên của các gia đình kênh natri và canxi phục vụ như là mục tiêu của thuốc giảm đau hiện đang được sử dụng, hoặc di truyền của con người hỗ trợ một vai trò trong việc truyền đau. Một đánh giá gần đây đã thảo luận về sự đóng góp quan trọng của kênh kali KCNQ, bao gồm cả lợi ích điều trị của việc tăng hoạt động kênh K + để điều trị đau dai dẳng (Brown và Passmore, 2009).
Kênh Natri có điện áp
Một loạt các kênh natri được thể hiện trong các tế bào thần kinh somatosensory, bao gồm các kênh nhạy cảm tetrodotoxin (TTX) Nav1.1, 1.6 và 1.7, và các kênh nhạy cảm TTX, Nav1.8 và 1.9. Trong những năm gần đây, sự đóng góp của Nav1.7 đã nhận được nhiều sự chú ý, vì hoạt động thay đổi của kênh này dẫn đến một loạt các rối loạn đau ở người (Cox et al., 2006; Dib-Hajj và cộng sự, 2008). Bệnh nhân bị đột biến chức năng trong gen này không thể phát hiện các kích thích độc hại, và kết quả là bị thương do thiếu phản xạ bảo vệ. Ngược lại, một số đột biến có chức năng trong Nav1.7 dẫn đến khả năng siêu nhạy của kênh và có liên quan đến hai rối loạn đau khác nhau ở người, đau cơ và rối loạn đau cực đoan kịch phát, cả hai đều gây cảm giác nóng dữ dội (Estacion et al ., 2008; Fertleman và cộng sự, 2006; Yang và cộng sự, 2004). Các nghiên cứu trên động vật đã chứng minh rằng Nav1.7 được điều chỉnh cao trong một loạt các mô hình đau viêm. Thật vậy, phân tích chuột thiếu Nav1.7 trong C nociceptors hỗ trợ một vai trò quan trọng cho kênh này trong quá mẫn cơ học và nhiệt sau viêm, và phản ứng cấp tính với kích thích cơ khí độc hại (Nassar et al., 2004). Hơi ngạc nhiên, đau do chấn thương thần kinh không bị thay đổi, cho thấy rằng các phân nhóm kênh natri khác biệt, hoặc một quần thể khác của các tác nhân biểu hiện Nav1.7, góp phần gây đau thần kinh (Nassar và cộng sự, 2005).
Kênh natri Nav1.8 cũng được thể hiện rõ ở hầu hết các nociceptors C. Cũng giống như các loại động vật loại bỏ Nav1.7, những thiếu hụt hiển thị Nav1.8 thiếu hụt nhạy cảm với nhiệt vô hại hoặc độc hại, hoặc áp lực vô hại; tuy nhiên, chúng hiển thị phản ứng suy yếu với các kích thích cơ học độc hại (Akopian et al., 1999). Nav1.8 cũng được yêu cầu để truyền các kích thích lạnh, vì những con chuột thiếu kênh này không nhạy cảm với lạnh trên một phạm vi nhiệt độ rộng (Zimmermann et al., 2007). Điều này là do Nav1.8 là duy nhất trong số các kênh natri nhạy cảm với điện áp ở chỗ nó không làm bất hoạt ở nhiệt độ thấp, làm cho nó trở thành máy phát điện tiềm năng hành động chủ yếu trong điều kiện lạnh.
Điều thú vị là những con chuột chuyển gen thiếu bộ phận biểu hiện tế bào thần kinh cảm giác Nav1.8, đã bị xóa bởi biểu hiện đích của độc tố bạch hầu A (Abrahamsen và cộng sự, 2008), hiển thị phản ứng suy yếu cho cả kích thích cơ học ngưỡng thấp và cao và lạnh. Ngoài ra, quá mẫn cảm cơ học và nhiệt trong các mô hình đau viêm bị suy yếu nghiêm trọng. Các kiểu hình vi phân của chuột thiếu các kênh Nav1.8 so với việc xóa các nơron biểu hiện Nav1.8 có lẽ phản ánh sự đóng góp của nhiều loại kênh phụ natri kênh điện áp để truyền các thông điệp đau.
Các kênh natri có điện áp là mục tiêu của thuốc gây tê cục bộ, làm nổi bật tiềm năng phát triển các thuốc giảm đau theo loại phụ. Nav1.7 là một mục tiêu đặc biệt thú vị để điều trị hội chứng đau viêm, một phần, bởi vì các nghiên cứu di truyền của con người cho thấy rằng chất ức chế Nav1.7 nên giảm đau mà không làm thay đổi các quá trình sinh lý cơ bản khác (xem ở trên). Một ứng dụng tiềm năng khác của thuốc chẹn kênh natri có thể điều trị quá mẫn cực với cảm lạnh, một tác dụng phụ bất lợi đặc biệt phiền hà của hóa trị liệu dựa trên bạch kim, chẳng hạn như oxaliplatin (Attal et al., 2009). Các chất đối kháng của Nav1.8 (hoặc TRPM8) có thể làm giảm hiện tượng này hoặc các dạng khác của chứng mất ngủ do cảm lạnh. Cuối cùng, các tiện ích tuyệt vời của thuốc ức chế tái hấp thu serotonin và norepinephrine để điều trị đau thần kinh có thể, trên thực tế, kết quả từ khả năng của họ để chặn các kênh natri điện áp gated (Dick et al., 2007).
Các kênh Canxi có điện áp
Một loạt các kênh canxi định mức điện thế được thể hiện trong các cơ quan thụ cảm. Các kênh canxi loại N-, P / Q- và T được chú ý nhiều nhất. Các kênh loại P / Q được thể hiện ở các đầu cuối tiếp hợp trong lớp II-IV của sừng lưng. Vai trò chính xác của chúng trong sự thụ thai không được giải quyết hoàn toàn. Tuy nhiên, đột biến trong các kênh này có liên quan đến chứng đau nửa đầu liệt nửa người có tính chất gia đình (de Vries et al., 2009). Các kênh canxi loại N và T cũng được biểu hiện bởi các sợi C và được điều chỉnh theo các trạng thái sinh lý bệnh, như trong các mô hình bệnh thần kinh do tiểu đường hoặc sau các dạng chấn thương thần kinh khác. Động vật thiếu Cav2.2 hoặc 3.2 cho thấy giảm nhạy cảm với các kích thích cơ học hoặc nhiệt sau khi bị viêm hoặc chấn thương thần kinh, tương ứng (Cao, 2006; Swayne và Bourinet, 2008; Zamponi và cộng sự, 2009; Messinger và cộng sự, 2009). Hơn nữa,? -Conotoxin GVIA, chất ngăn chặn các kênh loại N, được sử dụng trong da (dưới dạng ziconotide) để giảm đau do ung thư khó chữa (Rauck và cộng sự, 2009).
Tất cả các kênh canxi đều là protein truyền động cấu tạo bởi tiểu đơn vị? 1 lỗ rỗng và tiểu đơn vị điều chế? 2 ?,? 2? hoặc? 2 ?. 2? tiểu đơn vị điều chỉnh mật độ dòng điện và động học của quá trình kích hoạt và bất hoạt. Trong C nociceptors, các? 2? tiểu đơn vị được điều hòa mạnh mẽ sau chấn thương thần kinh và đóng một vai trò quan trọng trong quá mẫn và dị ứng do chấn thương (Luo và cộng sự, 2001). Thật vậy, tiểu đơn vị này là mục tiêu của nhóm thuốc chống co giật gabapentinoid, hiện đang được sử dụng rộng rãi để điều trị đau thần kinh (Davies và cộng sự, 2007).
Đau dai dẳng: Cơ chế ngoại vi
Đau dai dẳng liên quan đến chấn thương hoặc bệnh tật (như tiểu đường, viêm khớp, hoặc tăng trưởng khối u) có thể là kết quả của sự thay đổi trong tính chất của các dây thần kinh ngoại vi. Điều này có thể xảy ra do hậu quả của tổn thương dây thần kinh, dẫn đến tăng sự phát bắn tự phát hoặc thay đổi trong tính chất dẫn truyền hoặc dẫn truyền thần kinh của chúng. Trong thực tế, công dụng của thuốc tê tại chỗ và ngay cả hệ thống để điều trị các bệnh đau thần kinh khác nhau (như đau dây thần kinh sau xơ vữa) có thể phản ánh hành động của chúng trên các kênh natri tích lũy trong các sợi thần kinh bị thương.
Các Milieu hóa học của viêm
Sự nhạy cảm ngoại biên thường là kết quả của những thay đổi liên quan đến viêm trong môi trường hóa học của sợi thần kinh (McMahon et al., 2008). Do đó, tổn thương mô thường đi kèm với sự tích tụ các yếu tố nội sinh được giải phóng từ các tế bào nociceptors hoặc tế bào không thần kinh cư trú bên trong hoặc xâm nhập vào vùng bị thương (bao gồm tế bào mast, basophils, tiểu cầu, đại thực bào, bạch cầu trung tính, tế bào nội mô, tế bào sừng, và nguyên bào sợi). Chung. những yếu tố này, được gọi là “chất gây viêm”, đại diện cho một loạt các phân tử báo hiệu, bao gồm các chất dẫn truyền thần kinh, peptide (chất P, CGRP, bradykinin), eicosinoid và các chất béo liên quan (prostaglandin, thromboxanes, leukotrienes, endocannabinoids), neurotrophins, cytokines , và chemokine, cũng như protease ngoại bào và proton. Đáng chú ý, nociceptors thể hiện một hoặc nhiều thụ thể bề mặt tế bào có khả năng nhận biết và đáp ứng với từng tác nhân gây viêm hoặc pro-algesic (Hình 4). Các tương tác như vậy làm tăng khả năng kích thích của sợi thần kinh, do đó nâng cao độ nhạy cảm với nhiệt độ hoặc cảm ứng.
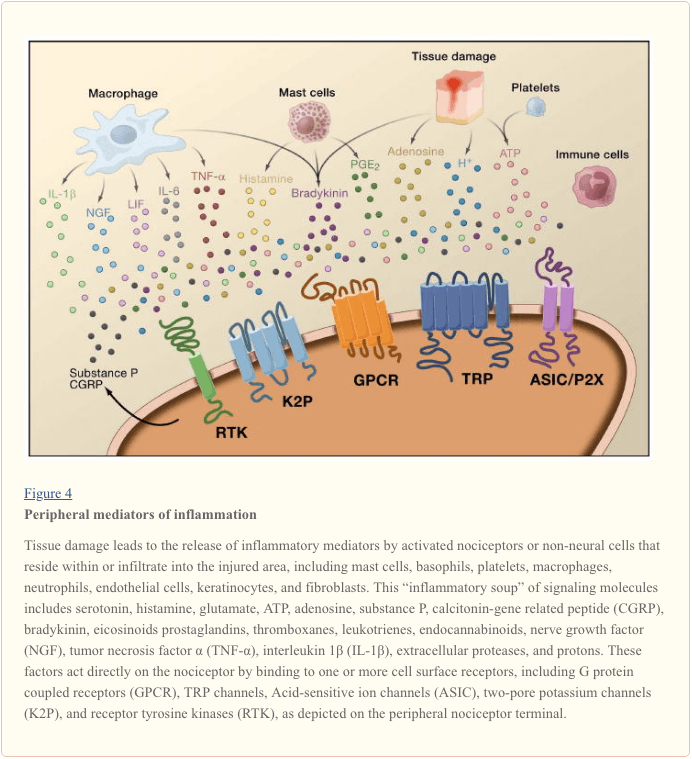
Không nghi ngờ gì là cách tiếp cận phổ biến nhất để giảm đau viêm liên quan đến việc ức chế sự tổng hợp hoặc tích lũy các thành phần của súp viêm. Điều này là tốt nhất được minh họa bởi các thuốc chống viêm không steroid, chẳng hạn như aspirin hoặc ibuprofen, làm giảm đau viêm và hyperalgesia bằng cách ức chế cyclooxygenases (Cox-1 và Cox-2) tham gia vào tổng hợp prostaglandin. Cách tiếp cận thứ hai là ngăn chặn các tác nhân của các tác nhân gây viêm tại nociceptor. Ở đây, chúng tôi nêu bật các ví dụ cung cấp thông tin chi tiết mới về cơ chế tế bào của nhạy cảm ngoại vi, hoặc hình thành cơ sở của các chiến lược điều trị mới để điều trị đau do viêm.
NGF có lẽ được biết đến nhiều nhất với vai trò là yếu tố thần kinh cần thiết cho sự tồn tại và phát triển của các tế bào thần kinh cảm giác trong quá trình tạo phôi, nhưng ở người lớn, NGF cũng được tạo ra trong việc thiết lập tổn thương mô và tạo thành một thành phần quan trọng của súp viêm (Ritner et al., 2009). Trong số nhiều mục tiêu di động của nó, NGF hoạt động trực tiếp trên các nociceptors sợi peptidergic C, thể hiện khả năng hấp thu cao của thụ thể NGF tyrosine kinase, TrkA, cũng như thụ thể thần kinh ái lực thấp, p75 (Chao, 2003; Snider và McMahon, 1998). NGF tạo ra quá mẫn sâu sắc đối với nhiệt và kích thích cơ học thông qua hai cơ chế phân biệt thời gian. Đầu tiên, một tương tác NGF-TrkA kích hoạt các đường truyền tín hiệu hạ lưu, bao gồm phospholipase C (PLC), protein kinase kích hoạt mitogen (MAPK), và phosphoinositide 3-kinase (PI3K). Điều này dẫn đến việc tăng cường chức năng của các protein mục tiêu tại đầu cuối nociceptor ngoại vi, đáng chú ý nhất là TRPV1, dẫn đến sự thay đổi nhanh chóng về độ nhạy nhiệt của tế bào và hành vi (Chuang et al., 2001). Ngoài những hành động nhanh này, NGF cũng được vận chuyển ngược đến hạt nhân của nociceptor, ở đó nó thúc đẩy sự biểu hiện của các protein chuyên sâu, bao gồm P, TRPV1 và tiểu đơn vị kênh natri điện áp Nav1.8 (Chao, 2003; Ji và cộng sự, 2002). Cùng với nhau, những thay đổi trong biểu hiện gen tăng cường kích thích của nociceptor và khuếch đại phản ứng viêm neurogenic.
Ngoài tế bào thần kinh, chấn thương thúc đẩy giải phóng nhiều cytokine, chủ yếu trong số đó là interleukin 1? (IL-1?) Và IL-6, và yếu tố hoại tử khối u? (TNF-?) (Ritner và cộng sự, 2009). Mặc dù có bằng chứng hỗ trợ tác động trực tiếp của các cytokine này lên các thụ thể, nhưng sự đóng góp chính của chúng vào quá mẫn cảm với cơn đau là kết quả của việc tăng phản ứng viêm và tăng sản xuất các tác nhân hỗ trợ giảm đau (chẳng hạn như prostaglandin, NGF, bradykinin và proton ngoại bào).
Bất kể cơ chế chống thụ cảm của chúng, việc can thiệp vào tín hiệu neurotrophin hoặc cytokine đã trở thành một chiến lược chính để kiểm soát bệnh viêm hoặc dẫn đến đau. Cách tiếp cận chính liên quan đến việc chặn NGF hoặc TNF-? tác dụng với một kháng thể trung hòa. Trong trường hợp của TNF- ?, điều này đã có hiệu quả rõ rệt trong điều trị nhiều bệnh tự miễn, bao gồm cả viêm khớp dạng thấp, dẫn đến giảm đáng kể sự phá hủy mô và chứng tăng kali huyết kèm theo (Atzeni và cộng sự, 2005). Bởi vì các hoạt động chính của NGF trên cơ quan thụ cảm ở người lớn xảy ra trong quá trình viêm nhiễm, nên ưu điểm của phương pháp này là chứng tăng tiết máu sẽ giảm mà không ảnh hưởng đến cảm giác đau bình thường. Thật vậy, các kháng thể kháng NGF hiện đang được thử nghiệm lâm sàng để điều trị các hội chứng đau do viêm (Hefti và cộng sự, 2006).
Mục tiêu của Súp viêm
TRPV1. Quá mẫn mạnh với nhiệt có thể phát triển với viêm hoặc sau khi tiêm các thành phần cụ thể của súp viêm (như bradykinin hoặc NGF). Thiếu sự nhạy cảm như vậy ở chuột thiếu TRPV1 cung cấp hỗ trợ di truyền cho ý tưởng rằng TRPV1 là một thành phần quan trọng của cơ chế mà qua đó tình trạng viêm tạo ra tăng cường nhiệt (Caterina và cộng sự, 2000; Davis và cộng sự, 2000). Thật vậy, các nghiên cứu in vitro đã chỉ ra rằng TRPV1 hoạt động như một chất tích hợp tín hiệu đa thức có độ nhạy nhiệt có thể được điều chế sâu sắc bởi các thành phần của súp viêm (Tominaga et al., 1998). Một số tác nhân gây viêm (ví dụ, các proton ngoại bào và lipid) hoạt động như các chất điều chỉnh allosteric trực tiếp dương tính của kênh, trong khi các chất khác (bradykinin, ATP và NGF) liên kết với các thụ thể của chúng trên những người chủ yếu và điều chỉnh TRPV1 thông qua hoạt hóa của nội bào hạ lưu Đường dẫn tín hiệu. Trong cả hai trường hợp, các tương tác này dẫn đến sự giảm sâu trong ngưỡng kích hoạt nhiệt của kênh, cũng như tăng cường độ đáp ứng ở nhiệt độ ngưỡng cực đại - tương đương sinh lý của chứng mất ngủ và tăng áp tương ứng.
Tuy nhiên, vẫn còn tranh cãi liên quan đến các cơ chế báo hiệu nội bào có trách nhiệm điều chế TRPV1 (Lumpkin và Caterina, 2007). Gợi nhớ các kênh TRP của tổ tiên trong mắt bay, nhiều kênh TRP của động vật có vú được kích hoạt hoặc điều hòa tích cực bởi sự phân cắt qua trung gian phospholipase C của màng phosphatidyl inositol 4,5 bisphosphate (PIP2). Tất nhiên, có nhiều hậu quả hạ lưu của hành động này, bao gồm sự sụt giảm màng PIP2, làm tăng nồng độ diacylglycerol và các chất chuyển hóa của nó, làm tăng lượng canxi trong tế bào, cũng như kích hoạt các protein kinaza. Trong trường hợp của TRPV1, hầu hết, nếu không phải tất cả, những con đường này đã được liên quan đến quá trình nhạy cảm và nó vẫn còn để được nhìn thấy có liên quan nhất đến quá mẫn nhiệt hành vi. Tuy nhiên, có một thỏa thuận rộng rãi rằng điều chế TRPV1 có liên quan đến chứng quá mẫn do tổn thương mô, đặc biệt là trong tình trạng viêm. Điều này sẽ bao gồm các tình trạng như bị cháy nắng, nhiễm trùng, viêm khớp dạng thấp hoặc viêm xương khớp và bệnh viêm mũi. Một ví dụ thú vị khác bao gồm đau từ ung thư xương (Honore và cộng sự, 2009), nơi sự phát triển khối u và phá hủy xương được đi kèm với toan mô cực kỳ mạnh mẽ, cũng như sản xuất cytokine, neurotrophins và prostaglandin.
TRPA1. Như đã mô tả ở trên, TRPA1 được kích hoạt bởi các hợp chất tạo thành cộng hóa trị với dư lượng cysteine. Ngoài các chất độc từ môi trường, điều này bao gồm các chất điện phân phản ứng thiol nội sinh được tạo ra trong quá trình tổn thương và viêm mô, hoặc do hậu quả của stress oxy hóa hoặc nitơ. Đứng đầu trong số các tác nhân như vậy là 4-hydroxy-2-nonenal và 15-deoxy-? 12,14-prostaglandin J2, cả hai đều là ?,? aldehyde không bão hòa được tạo ra thông qua quá trình peroxy hóa hoặc mất nước tự phát của sứ giả lipid thứ hai (Andersson và cộng sự, 2008; Cruz-Orengo và cộng sự, 2008; Materazzi và cộng sự, 2008; Trevisani và cộng sự, 2007). Các chất chủ vận TRPA1 nội sinh khác bao gồm axit nitrooleic, hydrogen peroxide và hydrogen sulfide. Ngoài các tác nhân tác động trực tiếp này, TRPA1 cũng được điều chế gián tiếp bởi các tác nhân tăng cường độ nhạy, chẳng hạn như bradykinin, hoạt động thông qua các thụ thể kết hợp với PLC. Thật vậy, những con chuột thiếu TRPA1 cho thấy giảm đáng kể phản ứng của tế bào và hành vi đối với tất cả các tác nhân này, cũng như giảm quá mẫn cảm nhiệt và cơ học do tổn thương mô (Bautista và cộng sự, 2006; Kwan và cộng sự, 2006). Cuối cùng, vì TRPA1 đóng một vai trò quan trọng trong các phản ứng gây viêm thần kinh và các phản ứng viêm khác đối với cả tác nhân nội sinh và độc tố môi trường dễ bay hơi, nên sự đóng góp của nó đối với chứng viêm đường thở, chẳng hạn như xảy ra trong bệnh hen suyễn, được đặc biệt quan tâm. Thật vậy, sự phong tỏa di truyền hoặc dược lý đối với TRPA1 làm giảm tình trạng viêm đường thở ở mô hình loài gặm nhấm bị hen suyễn do dị nguyên (Caceres và cộng sự, 2009).
ASIC. Như đã nói ở trên, các kênh ASIC là thành viên của gia đình DEG / ENaC được kích hoạt bởi sự axit hóa, và do đó đại diện cho một vị trí quan trọng khác cho hoạt động của các proton ngoại bào do hậu quả của tổn thương mô hoặc sự trao đổi chất. Các phân nhóm ASIC có thể hình thành một loạt các kênh đồng phân hoặc dị tính, mỗi kênh có độ nhạy pH và cấu hình biểu hiện riêng biệt. Các kênh chứa loại phụ ASIC3 được thể hiện cụ thể bởi những người khám phá và đặc biệt được đại diện tốt trong các sợi bảo vệ cơ xương và tim. Trong các mô này, sự trao đổi chất kỵ khí dẫn đến sự tích tụ axit lactic và proton, kích hoạt nociceptors để tạo ra đau cơ xương hoặc tim (Immke và McCleskey, 2001). Điều thú vị là các kênh có chứa ASIC3 mở để đáp ứng với sự giảm pH vừa phải (ví dụ 7.4 thành 7.0) xảy ra với thiếu máu cục bộ tim (Yagi et al., 2006). Axit lactic cũng làm tăng đáng kể sự kích thích của proton qua cơ chế liên quan đến việc thải canxi (Immke và McCleskey, 2003). Do đó, các kênh có chứa ASIC3 phát hiện và tích hợp các tín hiệu đặc biệt liên quan đến thiếu máu cơ và theo cách này, có chức năng khác biệt với các cảm biến axit khác trên nhân tố chính, như TRPV1 hoặc các loại phụ kênh ASIC khác.
Đau dai dẳng: Cơ chế trung ương
Nhạy cảm trung tâm đề cập đến quá trình thông qua đó một trạng thái hyperexcitability được thành lập trong hệ thống thần kinh trung ương, dẫn đến tăng cường xử lý các thông điệp (đau) nociceptive (Woolf, 1983). Mặc dù có nhiều cơ chế liên quan đến sự nhạy cảm trung tâm ở đây, chúng ta tập trung vào ba: sự thay đổi trong truyền dẫn thần kinh glutamatergic / quá mẫn cảm thụ NMDA, mất kiểm soát ức chế bổ (disinhibition) và tương tác thần kinh-glial (Hình 5).
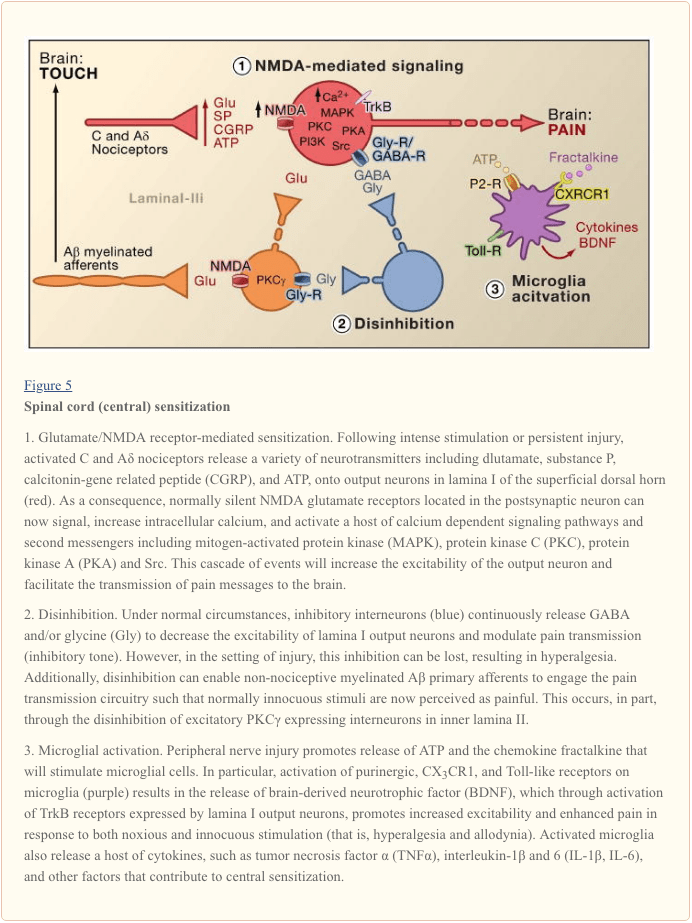
Nhạy cảm Glutamate / NMDA thụ thể qua trung gian
Đau cấp tính được báo hiệu bằng cách giải phóng glutamate từ các thiết bị đầu cuối trung tâm của nociceptors, tạo ra các dòng sau synapic kích thích (EPSC) trong các nơron sừng bậc hai. Điều này xảy ra chủ yếu thông qua hoạt hóa của AMPA sau synap và các phân nhóm kainate của các thụ thể glutamate ionotropic. Tóm tắt các ngưỡng EPSC phụ trong tế bào thần kinh sau synap cuối cùng sẽ dẫn đến khả năng hoạt động bắn và truyền thông điệp đau đến các nơron bậc cao. Trong các điều kiện này, phân loại NMDA của kênh glutamate là im lặng, nhưng trong thiết lập thương tích, sự phóng thích tăng chất dẫn truyền thần kinh từ nociceptors sẽ khử cực các neuron sau synap để kích hoạt các thụ thể NMDA không hoạt động. Sự gia tăng hậu quả của dòng canxi có thể tăng cường các kết nối khớp thần kinh giữa các nociceptors và các nơron truyền đau lưng sừng, do đó sẽ làm trầm trọng thêm các phản ứng với các kích thích độc hại (tức là tạo ra hyperalgesia).
Theo nhiều cách, quy trình này có thể so sánh với những thay đổi liên quan đến sự tăng cường dài hạn của vùng đồi thị (LTP) (để xem xét trên LTP trong lộ trình giảm đau, xem Drdla và Sandkuhler, 2008). Thật vậy, các loại thuốc làm tắc nghẽn cột sống LTP làm giảm chứng tăng áp do tổn thương mô. Như trong trường hợp LTP hippocampal, sự nhạy cảm trung tâm tủy sống phụ thuộc vào độ cao trung gian NMX của Ca2 + trong tế bào thần kinh sau synap. Kích hoạt đồng thời các glutamate metabotropic và các thụ thể P chất trên tế bào thần kinh sau synap cũng có thể góp phần gây nhạy cảm bằng cách làm tăng canxi cytosolic. Sự kích hoạt hạ lưu của một loạt các đường truyền tín hiệu và hệ thống tin nhắn thứ hai, đặc biệt là kinase (như MAPK, PKA, PKC, PI3K, Src), làm tăng khả năng kích thích của các nơron này, một phần bằng cách điều chỉnh chức năng thụ thể NMDA (Latremoliere và Woolf, 2009) . Minh họa của mô hình này là minh chứng rằng tiêm cột sống của một phân đoạn peptide chín axit amin của Src không chỉ làm gián đoạn sự tương tác thụ thể NMDI-Src mà còn làm giảm đáng kể quá mẫn do chấn thương ngoại vi gây ra mà không làm thay đổi cơn đau cấp tính. Src null chuột đột biến cũng hiển thị giảm allodynia cơ khí sau khi chấn thương dây thần kinh (Liu et al., 2008).
Ngoài việc tăng cường các yếu tố đầu vào từ vị trí chấn thương (chứng tăng tiết nguyên phát), sự nhạy cảm trung tâm góp phần vào tình trạng kích thích vô hại các khu vực xung quanh vị trí chấn thương có thể tạo ra đau. Chứng tăng phôtôn thứ phát này liên quan đến sự tạo điều kiện thuận lợi cho dị ứng, trong đó đầu vào từ A? những người hướng tâm, vốn thường phản ứng với chạm nhẹ, giờ tham gia vào các mạch dẫn truyền cảm giác đau, dẫn đến chứng rối loạn cơ học nghiêm trọng. Thực tế là khối chèn ép của sợi thần kinh ngoại vi đồng thời làm gián đoạn dẫn truyền ở A? hướng tâm và loại bỏ chứng tăng tiết thứ phát cho thấy rằng những mạch bất thường này được thiết lập trong các cơ sở lâm sàng cũng như trên các mô hình động vật (Campbell và cộng sự, 1988).
Mất kiểm soát GABAergic và Glycinergic: Disinhibition
GABAergic hoặc glycinergic ức chế interneurons được phân phối dày trong sừng lưng hời hợt và là cơ sở của lý thuyết kiểm soát cửa lâu dài của đau, mà postulates rằng mất chức năng của các interneurons ức chế (disinhibition) sẽ dẫn đến tăng đau (Melzack và Wall, 1965). Thật vậy, trong động vật gặm nhấm, quản trị cột sống của GABA (bicuculline) hoặc glycine (strychnine) đối kháng thụ thể (Malan et al., 2002; Sivilotti và Woolf, 1994; Yaksh, 1989) sản xuất quá mẫn hành vi tương tự như quan sát sau chấn thương ngoại vi. Phù hợp với những quan sát này, chấn thương ngoại vi dẫn đến sự suy giảm các dòng sau synap ức chế ở các nơron sừng phía trên bề mặt. Mặc dù Moore et al. (2002) cho rằng kết quả disinhibition do tử vong do tổn thương dây thần kinh ngoại vi gây ra tử cung GABAergic, tuyên bố này đã bị tranh chấp (Polgar et al., 2005). Bất kể nguyên nhân, sự ức chế thuốc giảm kết quả làm tăng sự khử cực và kích thích các nơron chiếu. Đối với sự nhạy cảm trung tâm NMDA, sự mất cân bằng tăng cường sản lượng tủy sống để đáp ứng với kích thích đau đớn và không đau đớn, góp phần gây ra chứng mất ngủ cơ học (Keller et al., 2007; Torsney và MacDermott, 2006).
Theo một báo cáo trước đó rằng việc xóa gen mã hóa PKC? ở chuột dẫn đến giảm rõ rệt chứng quá mẫn cảm cơ học do chấn thương thần kinh (Malmberg và cộng sự, 1997), các nghiên cứu gần đây đề cập đến sự tham gia của các tế bào thần kinh này trong quá trình khử trùng. Do đó, sau khi phong tỏa sự ức chế glycinergic bằng strychnine, việc đánh răng vô thưởng vô phạt sau sẽ kích hoạt các tế bào thần kinh cảm ứng PKC trong lamina II (Miraucourt và cộng sự, 2007), cũng như các tế bào thần kinh chiếu trong lamina I. Bởi vì tế bào thần kinh cảm ứng PKC? tủy sống chỉ nằm ở phần trong cùng của lamina II (Hình 1), theo đó những tế bào thần kinh này rất cần thiết cho sự biểu hiện của cơn đau dai dẳng do chấn thương dây thần kinh gây ra, và các cơ chế ức chế dẫn đến sự tăng hoạt của chúng.
Các nghiên cứu khác chỉ ra rằng chính những thay đổi trong tế bào thần kinh chiếu góp phần vào quá trình không ức chế. Ví dụ, tổn thương dây thần kinh ngoại vi điều chỉnh sâu sắc chất đồng vận chuyển K + -Cl- KCC2, chất cần thiết để duy trì gradien K + và Cl- bình thường qua màng sinh chất (Coull và cộng sự, 2003). Điều hòa KCC2, được biểu hiện ở các nơron chiếu lớp I, dẫn đến sự thay đổi gradient Cl-, như vậy sự hoạt hóa của các thụ thể GABA-A làm khử cực, thay vì siêu phân cực các nơron chiếu lớp I. Do đó, điều này sẽ tăng cường khả năng kích thích và tăng khả năng lây truyền cơn đau. Thật vậy, sự phong tỏa dược lý hoặc điều hòa giảm KCC2 qua trung gian siRNA ở chuột gây ra chứng dị ứng cơ học. Tuy nhiên, Zeilhofer và các đồng nghiệp cho rằng, ngay cả sau khi bị chấn thương, trương lực ức chế vẫn còn đủ để tăng cường dẫn truyền thần kinh GABAergic cột sống có thể là một cách tiếp cận có giá trị để giảm đau quá mẫn do chấn thương thần kinh ngoại vi (Knabl và cộng sự, 2008). Trên thực tế, các nghiên cứu trên chuột cho thấy rằng các loại thuốc nhắm mục tiêu cụ thể vào các phức hợp GABAA có chứa tiểu đơn vị? 2 và / hoặc? 3 làm giảm đau do viêm và đau thần kinh mà không tạo ra tác dụng phụ gây ngủ thường liên quan đến benzodiazepine, giúp tăng cường hoạt động của các kênh chứa? 1.
Disinhibition cũng có thể xảy ra thông qua điều chế tín hiệu glycinergic. Trong trường hợp này, cơ chế liên quan đến hoạt động cột sống của prostaglandin (Harvey và cộng sự, 2004). Cụ thể, tổn thương mô gây ra giải phóng cột sống của prostaglandin, PGE2, hoạt động trên các thụ thể EP2 được biểu hiện bởi các nội tiết kích thích và các nơron chiếu ở sừng lưng bề ngoài. Kích thích kết quả của đường dẫn cAMP-PKA phosphorylates các tiểu đơn vị thụ thể glycine GlyRa3, khiến các tế bào thần kinh không đáp ứng với các tác dụng ức chế của glycine. Theo đó, những con chuột thiếu gen GlyRa3 đã giảm nhiệt và quá mẫn cảm cơ học trong các mô hình tổn thương mô.
Tương tác Glial-Neuronal
Cuối cùng, các tế bào thần kinh đệm, đặc biệt là tế bào hình sao và tế bào hình sao, cũng góp phần vào quá trình nhạy cảm trung tâm xảy ra khi chấn thương. Trong điều kiện bình thường, microglia hoạt động như các đại thực bào thường trú của hệ thần kinh trung ương. Chúng phân bố đồng nhất trong chất xám của tủy sống và được cho là có chức năng như các trung đội thương tích hoặc nhiễm trùng. Tuy nhiên, trong vòng vài giờ sau khi bị chấn thương dây thần kinh ngoại vi, các microglia tích tụ ở sừng lưng bề ngoài trong vùng kết thúc của các sợi thần kinh ngoại vi bị thương. Microglia cũng bao quanh các thân tế bào của motoneurons sừng bụng, mà các sợi trục ngoại vi bị tổn thương đồng thời. Các microglia được kích hoạt giải phóng toàn bộ các phân tử tín hiệu, bao gồm các cytokine (chẳng hạn như TNF-?, Interleukin-1? Và 6), giúp tăng cường sự nhạy cảm trung tâm thần kinh và đau dai dẳng do chấn thương thần kinh (DeLeo và cộng sự, 2007). Thật vậy, tiêm microglia não đã hoạt hóa vào dịch não tủy ở mức tủy sống có thể tái tạo những thay đổi hành vi quan sát được sau chấn thương thần kinh (Coull và cộng sự, 2005). Do đó, dường như sự hoạt hóa vi mô là đủ để gây ra tình trạng đau dai dẳng (Tsuda và cộng sự, 2003).
Như microglia được kích hoạt sau dây thần kinh, nhưng không phải là tổn thương mô viêm, nó sau đó kích hoạt của xơ chất xơ, xảy ra trong cả hai điều kiện chấn thương, không phải là kích hoạt quan trọng cho kích hoạt microglial. Thay vào đó, thiệt hại vật chất của nhân viên ngoại vi phải tạo ra các tín hiệu cụ thể được phát hiện bởi microglia. Đứng đầu trong số này là ATP, nó nhắm vào các thụ thể purinergic loại P2 vi mô. Quan tâm đặc biệt là P2X4 (Tsuda et al., 2003), P2X7 (Chessell và cộng sự, 2005) và P2Y12 (Haynes et al., 2006; Kobayashi et al., 2008) kiểu phụ thụ thể. Thật vậy, ATP đã được sử dụng để kích hoạt microglia não trong các nghiên cứu cấy ghép tủy sống được đề cập ở trên (Tsuda et al., 2003). Hơn nữa, phong tỏa di truyền hoặc dược lý của chức năng thụ thể purinergic (Chessell và cộng sự, 2005; Tozaki-Saitoh và cộng sự, 2008; Ulmann và cộng sự, 2008) ngăn ngừa hoặc đảo ngược chứng mất ngủ cơ học do tổn thương thần kinh (Honore et al., 2006 ; Kobayashi và cộng sự, 2008; Tozaki-Saitoh và cộng sự, 2008; Tsuda và cộng sự, 2003).
Coull và các đồng nghiệp đã đề xuất một mô hình trong đó sự hoạt hóa microglia qua trung gian ATP / P2X4 gây ra cơ chế ức chế (Coull và cộng sự, 2005). Cụ thể, họ đã chứng minh rằng sự kích hoạt do ATP kích hoạt các thụ thể P2X4 gây ra sự giải phóng yếu tố dinh dưỡng thần kinh có nguồn gốc từ não (BDNF) từ microglia. Đến lượt mình, BDNF tác động lên các thụ thể TrkB trên các nơron chiếu lớp I, để tạo ra sự thay đổi trong gradient Cl-, như đã mô tả ở trên, sẽ chuyển hoạt động của GABA từ siêu phân cực sang khử cực. Liệu hiệu ứng do BDNF gây ra có liên quan đến biểu hiện KCC2, như xảy ra sau chấn thương thần kinh hay không, vẫn chưa được biết. Bất kể cơ chế nào, kết quả thực là sự kích hoạt microglia sẽ làm nhạy cảm các tế bào thần kinh lamina I sao cho phản ứng của chúng với đầu vào đơn âm từ các cơ quan thụ cảm hoặc đầu vào gián tiếp từ A? mối quan hệ, được nâng cao.
Ngoài BDNF, microglia hoạt hóa, giống như đại thực bào ngoại vi, giải phóng và phản ứng với nhiều chemokine và cytokine, và chúng cũng góp phần vào sự nhạy cảm trung tâm. Ví dụ, trong động vật không bị tổn thương (bình thường), chemokine fractalkine (CXCL1) được thể hiện bởi cả hai yếu tố chính và các tế bào thần kinh tủy sống (Lindia và cộng sự, 2005; Verge và cộng sự, 2004; Zhuang và cộng sự, 2007). Ngược lại, thụ thể fractalkine (CX3CR1) được biểu hiện trên các tế bào vi mô và quan trọng, được điều chỉnh sau chấn thương dây thần kinh ngoại vi (Lindia và cộng sự, 2005; Zhuang và cộng sự, 2007). Vì việc phân phối tủy xương có thể kích hoạt microglia, có vẻ như việc phát hành gãy xương do dây thần kinh gây ra do gãy xương cung cấp một tuyến đường khác thông qua đó vi khí hậu có thể được tham gia vào quá trình nhạy cảm trung tâm. Thật sự phong tỏa CX3CR1 với một kháng thể trung hòa ngăn cản cả sự phát triển và duy trì cơn đau dai dẳng gây thương tích (Milligan et al., 2004; Zhuang et al., 2007). Con đường này cũng có thể là một phần của một vòng phản hồi tích cực thông qua đó các sợi dây thần kinh bị tổn thương và các tế bào vi mô tương tác trong một thời gian tương hỗ và tái phát để khuếch đại tín hiệu đau. Điểm này được nhấn mạnh bởi thực tế là fractalkine phải được tách khỏi bề mặt thần kinh trước khi báo hiệu, một hành động được thực hiện bởi protease có nguồn gốc từ vi khuẩn, cathepsin S, các chất ức chế làm giảm allodynia gây ra tổn thương dây thần kinh và tăng áp (Clark et al., 2007). Quan trọng hơn, việc điều trị cột sống cathepsin S tạo ra quá mẫn cảm với hành vi ở dạng hoang dã, nhưng không ở chuột loại trực tiếp CX3CX1, liên kết cathepsin S với tín hiệu fractalkine (Clark và cộng sự, 2007; Zhuang và cộng sự, 2007). Mặc dù (các) yếu tố bắt đầu giải phóng cathepsin S từ microglia vẫn được xác định. ATP có vẻ là một khả năng hợp lý.
Gần đây, một số thành viên của họ thụ thể Toll giống như (TLRs) cũng đã được liên quan đến việc kích hoạt microglia sau chấn thương thần kinh. TLR là các protein truyền tín hiệu xuyên màng được biểu hiện trong các tế bào miễn dịch ngoại biên và glia. Là một phần của hệ miễn dịch bẩm sinh, chúng nhận ra các phân tử được chia sẻ rộng rãi bởi các tác nhân gây bệnh. Ức chế di truyền hoặc dược lý của TLR2, TLR3 hoặc TLR4 chức năng ở chuột kết quả không chỉ trong giảm kích hoạt microglial, mà còn làm giảm quá mẫn gây ra bởi chấn thương dây thần kinh ngoại biên (Kim et al., 2007; Obata et al., 2008; Tanga et al. , 2005). Không rõ là các phối tử nội sinh kích hoạt TLR2-4 sau chấn thương thần kinh. Trong số các ứng cử viên là mRNA hoặc protein sốc nhiệt có thể rò rỉ từ các tế bào thần kinh chủ yếu bị tổn thương và khuếch tán vào môi trường ngoại bào của tủy sống.
Sự đóng góp của các tế bào hình thành sự nhạy cảm trung tâm là ít rõ ràng hơn. Các tế bào hình cầu không thể nghi ngờ được gây ra trong tủy sống sau khi bị thương hoặc mô hoặc dây thần kinh (để xem xét, xem Ren và Dubner, 2008). Tuy nhiên, trái ngược với microglia, kích hoạt astrocyte thường bị trì hoãn và kéo dài lâu hơn, tới vài tháng. Một khả năng thú vị là các tế bào hình cầu quan trọng hơn đối với việc duy trì, hơn là cảm ứng sự nhạy cảm trung tâm và đau dai dẳng.
Cuối cùng, đáng chú ý là tổn thương ngoại biên không chỉ kích hoạt glia trong tủy sống, mà còn ở não, nơi glia đóng góp vào ảnh hưởng hỗ trợ siêu âm trong việc xử lý thông điệp đau trong tủy sống (xem Hình 2), hiện tượng có tên giảm dần thuận lợi (để xem xét, xem Ren và Dubner, 2008). Sự thuận lợi như vậy đặc biệt nổi bật trong bối cảnh chấn thương, và dường như chống lại các điều khiển ức chế phản hồi đồng thời phát sinh từ các loci não khác nhau (Porreca et al., 2002).

Sự hiểu biết của Tiến sĩ Alex Jimenez
Được thành lập bởi Hiệp hội Quốc tế về Nghiên cứu Đau hoặc IASP, nỗi đau là “trải nghiệm cảm xúc và cảm xúc khó chịu liên quan đến tổn thương mô hoặc mô tiềm năng, hoặc được mô tả về tổn thương mô hoặc cả hai. Nhiều nghiên cứu nghiên cứu đã được đề xuất để chứng minh cơ sở sinh lý của đau, tuy nhiên, không ai có thể bao gồm toàn bộ các khía cạnh liên quan đến nhận thức đau. Hiểu được cơ chế đau của cơn đau cấp tính so với đau mãn tính là cơ bản trong quá trình đánh giá lâm sàng vì điều này có thể giúp xác định phương pháp điều trị tốt nhất cho bệnh nhân có vấn đề sức khỏe tiềm ẩn.
Tính đặc hiệu trong việc truyền tải và kiểm soát thông điệp đau
Hiểu cách kích thích được mã hóa bởi hệ thần kinh để gợi ra những hành vi thích hợp là tầm quan trọng cơ bản đối với việc nghiên cứu tất cả các hệ thống giác quan. Trong hình thức đơn giản nhất, một hệ thống cảm giác sử dụng các dòng có nhãn để chuyển đổi các kích thích và gợi ra các hành vi thông qua các mạch được tách biệt hoàn toàn. Điều này có lẽ tốt nhất được chứng minh bởi hệ thống vị giác, nơi trao đổi một thụ thể ngọt ngào cho một người đắng trong một quần thể “các vị ngọt ngào” không làm thay đổi hành vi bị kích động bởi hoạt động trong dòng ghi nhãn đó; dưới những điều kiện này, một vị đắng cay đắng kích thích những người này để gợi ra một nhận thức về vị ngọt (Mueller et al., 2005).
Trong con đường đau đớn, cũng có bằng chứng để hỗ trợ sự tồn tại của các dòng nhãn. Như đã đề cập ở trên, nhiệt và lạnh được phát hiện bởi các tập con chủ yếu khác nhau của các sợi chủ yếu. Hơn nữa, việc loại bỏ các tập hợp con của người nuôi dưỡng có thể tạo ra thâm hụt có chọn lọc trong phản ứng hành vi với một phương thức độc hại cụ thể. Ví dụ, việc phá hủy các chất gây cảm ứng TRPV1 gây ra tổn thương nhiệt độ cao (bao gồm tăng cường nhiệt), không có sự thay đổi độ nhạy cảm với các kích thích cơ học hoặc cảm lạnh đau đớn. Ngược lại, việc xóa tập hợp con của MrgprD của nociceptors dẫn đến sự thâm hụt chọn lọc cao trong phản ứng cơ học, không có sự thay đổi về độ nhạy nhiệt (Cavanaugh et al., 2009). Bằng chứng thêm về sự phân biệt chức năng ở cấp độ của nociceptor xuất phát từ việc phân tích hai loại thụ thể opioid khác nhau (Scherrer và cộng sự, 2009). Cụ thể, các thụ thể opioid mu (MOR) chiếm ưu thế trong quần thể peptidergic, trong khi đó thụ thể opioid delta (DOR) được biểu hiện ở những người không phải là peptidergic nociceptors. Các chất chủ vận chọn lọc MOR ngăn chặn cơn đau nhiệt, trong khi các chất chủ vận chọn lọc DOR chặn đau cơ học, một lần nữa minh họa sự phân tách chức năng của quần thể nociceptor phân tử riêng biệt.
Những quan sát này tranh luận về tính đặc hiệu về mặt hành vi ở cấp độ của nociceptor. Tuy nhiên, đây có thể là một sự đơn giản hóa vì ít nhất hai lý do. Đầu tiên, nhiều nociceptors là polymodal và do đó có thể được kích hoạt bởi nhiệt, cơ khí, hoặc hóa học kích thích, để lại tự hỏi làm thế nào loại bỏ các nhóm lớn của nociceptors có thể có hiệu ứng cụ thể modality. Điều này lập luận cho một sự đóng góp đáng kể của mạch sống cho quá trình theo đó các tín hiệu nociceptive được mã hóa thành các phương thức đau khác nhau. Thật vậy, một mục tiêu quan trọng trong tương lai là phân định tốt hơn các phân nhóm thần kinh trong sừng lưng và mô tả tương tác khớp thần kinh của chúng với các quần thể được xác định về chức năng hoặc phân tử của người nuôi nociceptors. Thứ hai, hệ thống đau cho thấy khả năng thay đổi lớn, đặc biệt trong bối cảnh chấn thương, đặt ra câu hỏi về việc liệu hệ thống đường dây có dán nhãn có thể chứa được độ dẻo như thế hay không và thay đổi trong cơ chế đó như thế nào. Thật vậy, chúng ta biết rằng sự loại bỏ chất trung gian P-saporin của một số tế bào thần kinh sừng của I, tôi biểu hiện chất thụ thể P, có thể làm giảm cả quá mẫn cảm với nhiệt và cơ xảy ra sau mô hoặc tổn thương dây thần kinh (Nichols et al ., 1999). Những quan sát như vậy cho thấy rằng trong việc thiết lập độ đặc hiệu thương tích của dòng ghi nhãn không được duy trì nghiêm ngặt vì thông tin được truyền đến các mức cao hơn của chứng thần kinh.
Rõ ràng, các câu trả lời cho những câu hỏi này sẽ yêu cầu sử dụng kết hợp các phương pháp giải phẫu, điện sinh lý và hành vi để lập bản đồ mạch vật lý và chức năng làm nền tảng cho sự nociception và đau. Việc xác định liên tục các phân tử và gen đánh dấu các loại tế bào thần kinh cụ thể (cả ngoại vi và trung tâm) cung cấp các công cụ cần thiết để điều khiển các tế bào thần kinh di truyền hoặc dược lý và liên kết các hoạt động của chúng với các thành phần cụ thể của hành vi đau trong các hoàn cảnh bình thường và sinh lý bệnh. Làm như vậy sẽ đưa chúng ta đến gần hơn để tìm hiểu làm thế nào cơn đau cấp tính nhường đường cho những thay đổi không thích nghi gây ra đau mãn tính, và cách chuyển đổi này có thể được ngăn ngừa hoặc đảo ngược.

Hemp vs Marijuana: Sự khác biệt là gì?
Hiện nay, khoảng một nửa tiểu bang Hoa Kỳ cho phép bán cần sa y tế, và thậm chí một số thậm chí cho phép bán cần sa để sử dụng giải trí, càng ngày càng có nhiều người quan tâm đến các lợi ích sức khỏe có thể có của nhà máy gây tranh cãi này.
Trong khi khoa học về sử dụng y tế của nó vẫn tiếp tục tiến triển, nhiều người trong những ngày này đang cân nhắc cách họ có thể tiếp cận các lợi ích sức khỏe của cây mà không gặp phải tác dụng thần kinh không mong muốn nổi tiếng của nó. Điều này là hoàn toàn có thể với người thân của cần sa, cây gai dầu nhưng nó là điều cần thiết mà bạn nhận thức được sự khác biệt để bạn có thể là một người tiêu dùng thông minh.
Một cây trồng của cùng một nhà máy chính xác
Về cơ bản, cả cây gai dầu và cần sa là cùng một loại cây: Cannabis sativa. Có bằng chứng cho thấy Cannabis sativa L đã được trồng ở châu Á hàng ngàn năm về chất xơ cũng như nguồn cung cấp thực phẩm. Con người cuối cùng cũng nhận ra rằng ngọn hoa của cây có tính chất thần kinh. Với thời gian, khi con người đã làm như vậy với nhiều loại cây trồng khác, nông dân trồng cần sa bắt đầu trồng các loại cây cụ thể để tăng cường các đặc tính cụ thể.
Ngày nay, mặc dù một số người có thể tranh luận về số lượng thực sự của các loại thực vật, thực sự có hai sự phân biệt đơn giản,
Cây gai - Một loại cây trồng chủ yếu được trồng bên ngoài nước Mỹ, mặc dù một số nước Mỹ cho phép trồng trong mục đích nghiên cứu) để sử dụng trong quần áo, giấy, nhiên liệu sinh học, nhựa sinh học, thực phẩm bổ sung, mỹ phẩm và thực phẩm. Cây gai dầu được trồng ở ngoài trời như một loại cây trồng lớn với cả cây đực và cây con có mặt để thúc đẩy thụ phấn và cải thiện sản xuất hạt giống. Cây gai dầu công nghiệp nhập khẩu hợp pháp chứa ít hơn 0.3 phần trăm chất hóa học gây ung thư tetrahydrocannabinol, hoặc THC, nội dung của nó. Trong thực tế, cây gai dầu nhập khẩu hợp pháp thường sẽ loại bỏ cụ thể bất kỳ chất chiết xuất trong ngọn hoa khô của nhà máy.
Cần sa (Marihuana) - Cannabis sativa đặc biệt được trồng để tăng cường nội dung THC của nó được sử dụng cho mục đích y học hoặc giải trí. Cây cần sa thường được trồng trong nhà, trong điều kiện kiểm soát, và người trồng loại bỏ tất cả các cây đực từ vụ thu hoạch để ngăn thụ tinh vì phân bón làm giảm mức độ THC của cây.
Tính hợp pháp của cần sa y tế
Việc sử dụng cần sa y tế là một khu vực gia tăng tranh cãi cho các nhà nghiên cứu và người tiêu dùng. Mặc dù có thể không hoàn toàn một nửa các tiểu bang của Hoa Kỳ đã hợp pháp hóa việc sử dụng cây này, nó vẫn còn bất hợp pháp theo luật liên bang, và do đó việc sử dụng nó vẫn còn gây tranh cãi bất kể thực tế là dường như có lợi ích sức khỏe thực sự cho nhiều vấn đề sức khỏe nghiêm trọng.
Những người đang tìm cách sử dụng cần sa để sử dụng y tế nên nói về lợi ích của nó so với mối nguy hiểm của nó với một chuyên gia chăm sóc sức khỏe có tay nghề cao trước khi sử dụng nó. Ngoài ra, nhiều người tiêu dùng có quan tâm đến lợi ích sức khỏe của nó không cần các tác dụng phụ thần kinh của THC hoặc nguy cơ của một thử nghiệm thuốc dương tính.
Cây gai dầu: Lợi ích sức khỏe không có rủi ro
Cây gai dầu nhập khẩu, có mức THC rất thấp, gần như vắng mặt, có thể là giải pháp cho người tiêu dùng đang tìm kiếm lợi ích sức khỏe của cây trừ đi những ảnh hưởng của THC.
Mặc dù THC có một số lợi ích sức khỏe, gai bao gồm nhiều hợp chất hoạt tính sinh học 80 có thể cung cấp hỗ trợ tuyệt vời cho một loạt các vấn đề sức khỏe, như phản ứng căng thẳng, tâm trạng tích cực, và khó chịu về thể chất hoặc đau. Cây gai dầu cũng có thể có lợi cho sức khỏe đường tiêu hóa, giúp giữ một phản ứng viêm khỏe mạnh trong toàn bộ cơ thể, và hỗ trợ chức năng miễn dịch bình thường.
Nếu bạn đang xem xét việc sử dụng một sản phẩm bổ sung dinh dưỡng bao gồm cây gai dầu, thì đó là lý tưởng để mua một sản phẩm từ một nguồn đáng tin cậy.
Trong kết luận, cả hệ thống thần kinh ngoại vi và trung ương phát hiện, giải thích và điều chỉnh một loạt các kích thích nhiệt và cơ học cũng như các chất kích thích hóa học nội sinh và môi trường. Nếu các kích thích quá mạnh, nó có thể tạo ra cơn đau cấp tính, trong trường hợp đau dai dẳng hoặc mãn tính, sự truyền đau có thể bị ảnh hưởng rất nhiều. Bài báo trên mô tả cơ chế tế bào và phân tử của cơn đau để được hướng dẫn trong các đánh giá lâm sàng. Hơn nữa, việc sử dụng cây gai dầu có thể mang lại nhiều lợi ích cho sức khỏe so với những tác dụng gây tranh cãi của cần sa. Thông tin tham khảo từ Trung tâm Thông tin Công nghệ Sinh học Quốc gia (NCBI). Phạm vi thông tin của chúng tôi được giới hạn trong trị liệu thần kinh cột sống cũng như các chấn thương và tình trạng cột sống. Để thảo luận về chủ đề này, vui lòng hỏi Tiến sĩ Jimenez hoặc liên hệ với chúng tôi tại 915-850-0900 .
Quản lý bởi Tiến sĩ Alex Jimenez
Abrahamsen B, Zhao J, Asante CO, Cendan CM, Marsh S, Martinez-Barbera JP, Nassar MA, Dickenson AH, Gỗ JN. Các tế bào và cơ sở phân tử của đau cơ, lạnh và viêm. Khoa học. 2008; 321: 702 – 705. [PubMed]
Akopian AN, Souslova V, Anh S, Okuse K, Ogata N, Ure J, Smith A, Kerr BJ, McMahon SB, Boyce S, et al. Kênh natri kháng tetrodotoxin SNS có chức năng chuyên biệt trong các con đường đau. Nat Neurosci. 1999; 2: 541 – 548. [PubMed]
Alessandri-Haber N, Dina OA, Joseph EK, Reichling D, Levine JD. Một thụ thể thoáng qua tiềm năng vanilloid cơ chế phụ thuộc vào 4 của hyperalgesia được tham gia bởi hành động phối hợp của các chất trung gian gây viêm. J Neurosci. 2006; 26: 3864 – 3874. [PubMed]
Andersson DA, Gentry C, Moss S, Bevan S. Khả năng thụ thể thoáng qua A1 là một thụ thể cảm quan cho nhiều sản phẩm của stress oxy hóa. J Neurosci. 2008; 28: 2485 – 2494. [PMC bài viết miễn phí] [PubMed]
Apkarian AV, Bushnell MC, Treede RD, Zubieta JK. Cơ chế não bộ của con người về nhận thức đau và quy định về sức khỏe và bệnh tật. Eur J Pain. 2005; 9: 463 – 484. [PubMed]
Attal N, Bouhassira D, Gautron M, Vaillant JN, Mitry E, Lepere C, Rougier P, Guirimand F. Tăng cường nhiệt như là một dấu hiệu của độc tính thần kinh oxaliplatin: một nghiên cứu đánh giá cảm quan định lượng tương lai. Đau đớn. 2009; 144: 245 – 252. [PubMed]
Atzeni F, Turiel M, Capsoni F, Doria A, Meroni P, Sarzi-Puttini P. Các tác nhân tự miễn dịch và kháng TNF-alpha. Ann NY Acad Sci. 2005; 1051: 559 – 569. [PubMed]
Bandell M, Story GM, Hwang SW, Viswanath V, Eid SR, Petrus MJ, Earley TJ, Patapoutian A. Kênh ion lạnh độc hại TRPA1 được kích hoạt bởi các hợp chất hăng và bradykinin. Neuron. 2004; 41: 849 – 857. [PubMed]
Basbaum AI, Jessell T. Nhận thức về đau đớn. Trong: Kandel ER, Schwartz J, Jessell T, biên tập viên. Nguyên tắc của Neuroscience. New York: Appleton và Lange; 2000. pp. 472 – 491.
Bautista DM, Jordt SE, Nikai T, Tsuruda PR, Đọc AJ, Poblete J, Yamoah EN, AI Basbaum, Julius D. TRPA1 làm trung gian cho các tác nhân gây viêm của các chất kích thích môi trường và các tác nhân gây dị ứng. Ô. 2006; 124: 1269 – 1282. [PubMed]
Bautista DM, Siemens J, Glazer JM, Tsuruda PR, Basbaum AI, Stucky CL, Jordt SE, Julius D. Các thụ thể menthol TRPM8 là máy phát hiện chính của môi trường lạnh. Thiên nhiên. 2007; 448: 204 – 208. [PubMed]
Bautista DM, Sigal YM, Milstein AD, Garrison JL, Zorn JA, Tsuruda PR, Nicoll RA, Julius D. Các tác nhân hăng từ hạt Szechuan kích thích các tế bào thần kinh cảm giác bằng cách ức chế các kênh kali hai lỗ. Nat Neurosci. 2008; 11: 772 – 779. [PMC bài viết miễn phí] [PubMed]
Bessac BF, Jordt SE. Các kênh TRP hấp dẫn: TRPA1 và TRPV1 trong điều khiển phản xạ và phản xạ đường hô hấp. Sinh lý học (Bethesda) 2008; 23: 360 – 370. [PMC bài viết miễn phí] [PubMed]
Bhattacharya MR, Bautista DM, Wu K, Haeberle H, Lumpkin EA, Julius D. Radial stretch cho thấy các quần thể riêng biệt của các tế bào thần kinh somatosensory mammosensitive mammalian. Proc Natl Acad Sci Mỹ A. 2008; 105: 20015 – 20020. [PMC bài viết miễn phí] [PubMed]
Brown DA, Passmore GM. Neural KCNQ (Kv7) kênh. Br J Pharmacol. 2009; 156: 1185 – 1195. [PMC bài viết miễn phí] [PubMed]
Bryant BP, Mezine I. Các alkylamides tạo ra dị cảm ngứa ran kích hoạt các tế bào thần kinh sinh ba xúc giác và nhiệt. Brain Res. 1999; 842: 452 – 460. [PubMed]
Caceres AI, Brackmann M, Elia MD, Bessac BF, del Camino D, D'Amours M, Witek JS, Fanger CM, Chong JA, Hayward NJ, và cộng sự. Một kênh ion thần kinh cảm giác cần thiết cho viêm đường hô hấp và hyperreactivity trong hen suyễn. Proc Natl Acad Sci Mỹ A. 2009; 106: 9099 – 9104. [PMC bài viết miễn phí] [PubMed]
Campbell JN, Raja SN, Meyer RA, Mackinnon SE. Các bệnh nhân myelinated báo hiệu hyperalgesia kết hợp với chấn thương thần kinh. Đau đớn. 1988; 32: 89 – 94. [PubMed]
Cao YQ. Các kênh canxi có điện áp và đau. Đau đớn. 2006; 126: 5 – 9. [PubMed]
Caterina MJ, Leffler A, Malmberg AB, Martin WJ, Trafton J, Petersen-Zeitz KR, Koltzenburg M, Basbaum AI, Julius D. Khử độc giác và cảm giác đau ở chuột thiếu thụ thể capsaicin. Khoa học. 2000; 288: 306 – 313. [PubMed]
Caterina MJ, Rosen TA, Tominaga M, AJ phanh, Julius D. Một homologue capsaicin-thụ thể với một ngưỡng cao cho nhiệt độc hại. Thiên nhiên. 1999; 398: 436 – 441. [PubMed]
Caterina MJ, Schumacher MA, Tominaga M, Rosen TA, Levine JD, Julius D. Các thụ thể capsaicin: một kênh ion kích hoạt nhiệt trong con đường đau. Thiên nhiên. 1997; 389: 816 – 824. [PubMed]
Cavanaugh DJ, Lee H, Lo L, Shields SD, Zylka MJ, Basbaum AI, Anderson DJ. Các tập con riêng biệt của các sợi cảm giác chính không được kết hợp trung gian sẽ làm trung gian các phản ứng hành vi với các kích thích nhiệt và cơ khí độc hại. Proc Natl Acad Sci Mỹ A. 2009; 106: 9075 – 9080. [PMC bài viết miễn phí] [PubMed]
Cesare P, McNaughton P. Một hiện tượng mới kích hoạt nhiệt trong các tế bào thần kinh nociceptive và sự nhạy cảm của nó bởi bradykinin. Proc Natl Acad Sci Mỹ A. 1996; 93: 15435 – 15439. [PMC bài viết miễn phí] [PubMed]
Chalfie M. Neurosensory mechanotransduction. Nat Rev Mol Cell Biol. 2009; 10: 44 – 52. [PubMed]
Chao MV. Neurotrophins và thụ thể của chúng: một điểm hội tụ cho nhiều con đường tín hiệu. Nat Rev Neurosci. 2003; 4: 299 – 309. [PubMed]
Chen L, Huang LY. Protein kinase C làm giảm Mg2 + khối của các kênh thụ thể NMDA như một cơ chế điều chế. Thiên nhiên. 1992; 356: 521 – 523. [PubMed]
Chen X, Alessandri-Haber N, Levine JD. Đánh dấu sự suy giảm của sự nhạy cảm với chất gây xơ C gây ra bởi chất kích thích trung gian cho các kích thích cơ học và hạ huyết áp ở chuột TRPV4 - / -. Đau Mol. 2007; 3: 31. [PMC bài viết miễn phí] [PubMed]
IP của Chessell, Hatcher JP, Bountra C, Michel AD, Hughes JP, Green P, Egerton J, Murfin M, Richardson J, Peck WL, et al. Sự phá vỡ gen purinoceptor của P2X7 làm giảm đau mãn tính và đau thần kinh. Đau đớn. 2005; 114: 386 – 396. [PubMed]
Cho H, Koo JY, Kim S, Park SP, Yang Y, Oh U. Một kênh nhạy cảm cơ học mới được xác định trong các tế bào thần kinh cảm giác. Eur J Neurosci. 2006; 23: 2543 – 2550. [PubMed]
Cho H, Shin J, Shin CY, Lee SY, Oh U. Các kênh ion cơ học trong tế bào thần kinh cảm giác nuôi cấy của chuột sơ sinh. J Neurosci. 2002; 22: 1238 – 1247. [PubMed]
Chuang HH, Prescott ED, Kong H, Shields S, Jordt SE, Basbaum AI, Chao MV, Julius D. Bradykinin và yếu tố tăng trưởng thần kinh giải phóng thụ thể capsaicin từ ức chế trung gian P4,5 của PtdIns (2). Thiên nhiên. 2001; 411: 957 – 962. [PubMed]
Chung MK, Lee H, Caterina MJ. Nhiệt độ ấm kích hoạt TRPV4 trong chuột 308 keratinocytes. J Biol Chem. 2003; 278: 32037 – 32046. [PubMed]
Clark AK, Yip PK, Grist J, Gentry C, Staniland AA, Marchand F, Dehvari M, Wotherspoon G, Mùa đông J, Ullah J, et al. Ức chế cathepsin vi mô cột sống S để đảo ngược đau thần kinh. Proc Natl Acad Sci Mỹ A. 2007; 104: 10655 – 10660. [PMC bài viết miễn phí] [PubMed]
Colburn RW, Lubin ML, Stone DJ, Jr, Wang Y, Lawrence D, D'Andrea MR, Brandt MR, Liu Y, Flores CM, Tần N. Cảm nhận lạnh nhạy cảm với chuột TRPM8. Neuron. 2007; 54: 379 – 386. [PubMed]
Bài JA, Beggs S, Boudreau D, Boivin D, Tsuda M, Inoue K, Gravel C, Salter MW, De Koninck Y. BDNF từ microglia gây ra sự thay đổi trong anion xoắn thần kinh nằm dưới đau thần kinh. Thiên nhiên. 2005; 438: 1017 – 1021. [PubMed]
Tác giả JA, Boudreau D, Bachand K, Prescott SA, Nault F, Sik A, De Koninck P, De Koninck Y. Chuyển dịch khớp thần kinh trong anion gradient trong các tế bào thần kinh cột sống I như một cơ chế của đau thần kinh. Thiên nhiên. 2003; 424: 938 – 942. [PubMed]
Cox JJ, Reimann F, Nicholas AK, Thornton G, Roberts E, Springell K, Karbani G, Jafri H, Mannan J, Raashid Y, et al. Bệnh mạch máu SCN9A gây khó chịu bẩm sinh để cảm thấy đau. Thiên nhiên. 2006; 444: 894 – 898. [PubMed]
Cruz-Orengo L, Dhaka A, Heuermann RJ, Young TJ, Montana MC, Cavanaugh EJ, Kim D, Story GM. Nhiễm niệu da gây ra bởi 15-delta PGJ2 thông qua kích hoạt kênh ion TRPA1. Đau Mol. 2008; 4: 30. [PMC bài viết miễn phí] [PubMed]
Davies A, Hendrich J, Văn Minh AT, Wratten J, Douglas L, Dolphin AC. Sinh học chức năng của các tiểu đơn vị delta alpha (2) của các kênh canxi có điện áp. Xu hướng Pharmacol Sci. 2007; 28: 220 – 228. [PubMed]
Davis JB, Gray J, Gunthorpe MJ, Hatcher JP, Davey PT, Overend P, Harries MH, Latcham J, Clapham C, Atkinson K, et al. Thụ thể Vanilloid-1 là điều cần thiết cho tăng cường nhiệt viêm. Thiên nhiên. 2000; 405: 183 – 187. [PubMed]
DeLeo JA, Sorkin LS, Watkins LR, biên tập viên. Điều trị đau và miễn dịch. IASP; Seattle: 2007.
de Vries B, Frants RR, Ferrari MD, van den Maagdenberg AM. Di truyền phân tử của chứng đau nửa đầu. Hum Genet. 2009; 126: 115 – 132. [PubMed]
Dhaka A, Murray AN, Mathur J, Earley TJ, Petrus MJ, Patapoutian A. TRPM8 là cần thiết cho cảm giác lạnh ở chuột. Neuron. 2007; 54: 371 – 378. [PubMed]
Dib-Hajj SD, Yang Y, Waxman SG. Di truyền và sinh lý bệnh phân tử của Na (v) hội chứng đau liên quan đến 1.7. Adv Genet. 2008; 63: 85 – 110. [PubMed]
Dick IE, Brochu RM, Purohit Y, Kaczorowski GJ, Martin WJ, Linh mục BT. Sự phong tỏa kênh natri có thể góp phần vào hiệu quả giảm đau của thuốc chống trầm cảm. J Đau. 2007; 8: 315 – 324. [PubMed]
Dobler TM, Springauf A, Tovornik S, Weber M, Schmitt A, Sedlmeier R, Wischmeyer E, Doring F. TRESK các kênh K + hai lỗ rỗng tạo thành một thành phần quan trọng của các dòng kali nền trong các tế bào thần kinh DRG chuột. J Physiol 2007 [PMC bài viết miễn phí] [PubMed]
Dong X, Han S, Zylka MJ, Simon MI, Anderson DJ. Một nhóm gia đình GPCR đa dạng thể hiện trong các tập con cụ thể của các tế bào thần kinh cảm giác nociceptive. Ô. 2001; 106: 619 – 632. [PubMed]
Drdla R, Sandkuhler J. Sự tăng cường dài hạn ở các khớp thần kinh C-xơ nhờ hoạt động ức chế ở mức độ thấp trong cơ thể. Đau Mol. 2008; 4: 18. [PMC bài viết miễn phí] [PubMed]
Drew LJ, Rohrer DK, MP giá, Blaver KE, Cockayne DA, Cesare P, Gỗ JN. Các kênh ion cảm biến axit ASIC2 và ASIC3 không đóng góp vào các dòng hoạt hóa cơ học trong các tế bào thần kinh cảm giác của động vật có vú. J Physiol. 2004; 556: 691 – 710. [PMC bài viết miễn phí] [PubMed]
Drew LJ, Wood JN, Cesare P. Các đặc tính nhạy cảm về cơ học của các nơron cảm giác capsaicin nhạy cảm và không nhạy cảm. J Neurosci. 2002; 22: RC228. [PubMed]
Estacion M, Dib-Hajj SD, Benke PJ, Te Morsche RH, Eastman EM, Macala LJ, JP thứ mười, Waxman SG. NaV1.7 tăng đột biến chức năng như là một liên tục: A1632E hiển thị các thay đổi sinh lý liên quan đến đau hồng cầu và đột biến rối loạn cơn đau do kịch phát và gây ra các triệu chứng của cả hai rối loạn. J Neurosci. 2008; 28: 11079 – 11088. [PubMed]
Fertleman CR, Baker MD, Parker KA, Moffatt S, Elmslie FV, Abrahamsen B, Ostman J, Klugbauer N, Gỗ JN, Gardiner RM, et al. Đột biến SCN9A trong rối loạn đau cực đoan kịch phát: các biến thể allelic phụ thuộc vào dị tật kênh và kiểu hình riêng biệt. Neuron. 2006; 52: 767 – 774. [PubMed]
Gevaert T, Vriens J, Segal A, Everaerts W, Roskams T, Talavera K, Owsianik G, Liedtke W, Daelemans D, Dewachter I, et al. Việc xóa kênh cation tiềm năng thụ thể thoáng qua TRPV4 làm suy yếu việc làm bàng quang bàng quang. J Clin Invest. 2007; 117: 3453 – 3462. [PMC bài viết miễn phí] [PubMed]
Cấp AD, Cottrell GS, Amadesi S, Trevisani M, Nicoletti P, Materazzi S, Altier C, Cenac N, Zamponi GW, Bautista-Cruz F, et al. Thụ thể protease kích hoạt 2 nhạy cảm với các thụ thể thoáng qua tiềm năng vanilloid kênh ion 4 gây ra tăng áp cơ học ở chuột. J Physiol. 2007; 578: 715 – 733. [PMC bài viết miễn phí] [PubMed]
Guler AD, Lee H, Iida T, Shimizu I, Tominaga M, Caterina M. Kích hoạt nhiệt của kênh ion, TRPV4. J Neurosci. 2002; 22: 6408 – 6414. [PubMed]
Harvey RJ, Người phụ trách UB, Wassle H, Ahmadi S, Heindl C, Reinold H, Thông minh TG, Harvey K, Schutz B, Abo-Salem OM, và cộng sự. GlyR alpha3: một mục tiêu thiết yếu đối với sự nhạy cảm đau do PGE2 qua trung gian cột sống. Khoa học. 2004; 304: 884 – 887. [PubMed]
Haynes SE, Hollopeter G, Yang G, Kurpius D, Dailey ME, Gan WB, Julius D. Các thụ thể P2Y12 điều chỉnh kích hoạt microglial bởi các nucleotide ngoại bào. Nat Neurosci. 2006; 9: 1512 – 1519. [PubMed]
Hensel H, Zotterman Y. Tác dụng của tinh dầu bạc hà trên các thermoreceptors. Acta Physiol Scand. 1951; 24: 27 – 34. [PubMed]
Hefti FF, Rosenthal A, Walicke PA, Wyatt S, Vergara G, Shelton DL, Davies AM. Tiểu thuyết loại thuốc giảm đau dựa trên tính đối kháng của NGF. Xu hướng Pharmacol Sci. 2006; 27: 85 – 91. [PubMed]
Hensel H, Zotterman Y. Tác dụng của tinh dầu bạc hà trên các thermoreceptors. Acta Physiol Scand. 1951; 24: 27 – 34. [PubMed]
Hill K, Schaefer M. TRPA1 được điều chế khác biệt bởi các phân tử lưỡng tính trinitrophenol và chlorpromazine. J Biol Chem. 2007; 282: 7145 – 7153. [PubMed]
Hinman A, Chuang HH, Bautista DM, Julius D. TRP kênh kích hoạt bằng cách sửa đổi cộng hóa trị nghịch đảo. Proc Natl Acad Sci Mỹ A. 2006; 103: 19564 – 19568. [PMC bài viết miễn phí] [PubMed]
Honore P, Chandran P, Hernandez G, Gauvin DM, Mikusa JP, Zhong C, Joshi SK, Ghilardi JR, Sevcik MA, Fryer RM, et al. Liều lặp đi lặp lại của ABT-102, một chất đối kháng TRPV1 mạnh và có chọn lọc, tăng cường hoạt động giảm đau qua trung gian TRPV1 ở động vật gặm nhấm, nhưng làm giảm sự tăng thân nhiệt đối kháng. Đau đớn. 2009; 142: 27 – 35. [PubMed]
Honore P, Donnelly-Roberts D, Namovic MT, Hsieh G, Zhu CZ, Mikusa JP, Hernandez G, Trung C, Gauvin DM, Chandran P, et al. A-740003 [N- (1 - {[(cyanoimino) (5-quinolinylamino) metyl] amino} -2,2-dimethylpropyl) -2- (3,4-dimethoxyphenyl) acetamide], một chất đối kháng thụ thể P2X7 mới và có chọn lọc, liều phụ thuộc giảm đau thần kinh ở chuột. J Pharmacol Exp Ther. 2006; 319: 1376 – 1385. [PubMed]
Hu J, Lewin GR. Các dòng điện cơ học trong các tế bào thần kinh của các tế bào thần kinh cảm giác chuột nuôi cấy. J Physiol. 2006; 577: 815 – 828. [PMC bài viết miễn phí] [PubMed]
Ngưng DC, McCleskey EW. Lactate tăng cường kênh cảm ứng axit Na + trên các tế bào thần kinh cảm biến thiếu máu cục bộ. Nat Neurosci. 2001; 4: 869 – 870. [PubMed]
Ngưng DC, McCleskey EW. Proton mở các kênh ion cảm biến axit bằng cách xúc tác cứu trợ Ca2 + phong tỏa. Neuron. 2003; 37: 75 – 84. [PubMed]
Ji RR, Samad TA, Jin SX, Schmoll R, Woolf CJ. kích hoạt P38 MAPK bởi NGF trong các tế bào thần kinh cảm giác chính sau khi viêm làm tăng mức TRPV1 và duy trì tăng cường nhiệt. Neuron. 2002; 36: 57 – 68. [PubMed]
Jordt SE, Bautista DM, Chuang HH, McKemy DD, Zygmunt PM, Hogestatt ED, Meng ID, dầu Julius D. Mustard và cannabinoids kích thích các dây thần kinh cảm giác thông qua kênh TRP ANKTM1. Thiên nhiên. 2004; 427: 260 – 265. Epub 2004 Jan 2007. [PubMed]
Julius D, Basbaum AI. Cơ chế phân tử của nociception. Thiên nhiên. 2001; 413: 203 – 210. [PubMed]
Karashima Y, Damann N, Prenen J, Talavera K, Segal A, Voets T, Nilius B. Hành động Bimodal của tinh dầu bạc hà trên kênh tiềm năng thụ thể thoáng qua TRPA1. J Neurosci. 2007; 27: 9874 – 9884. [PubMed]
Karashima Y, Talavera K, Everaerts W, Janssens A, Kwan KY, Vennekens R, Nilius B, Voets T. TRPA1 hoạt động như một cảm biến lạnh trong ống nghiệm và in vivo. Proc Natl Acad Sci Mỹ A. 2009; 106: 1273 – 1278. [PMC bài viết miễn phí] [PubMed]
Kawasaki Y, Kohno T, Zhuang ZY, Brenner GJ, Wang H, Van Der Meer C, Befort K, Woolf CJ, Ji RR. Các thụ thể ionotropic và metabotropic, protein kinase A, protein kinase C và Src đóng góp vào hoạt hóa ERK của sợi C và phản ứng phosphoryl hóa protein liên kết yếu tố trong tế bào sừng lưng, dẫn đến sự nhạy cảm trung tâm. J Neurosci. 2004; 24: 8310 – 8321. [PubMed]
Keller AF, Beggs S, Salter MW, De Koninck Y. Chuyển đổi sản lượng của các tế bào thần kinh cột sống I sau tổn thương dây thần kinh và kích thích microglia nằm dưới đau thần kinh. Đau Mol. 2007; 3: 27. [PMC bài viết miễn phí] [PubMed]
Kim D, Kim MA, Cho IH, Kim MS, Lee S, Jo EK, Choi SY, Park K, Kim JS, Akira S, et al. Một vai trò quan trọng của thụ thể giống như 2 trong kích hoạt tế bào thần kinh cột sống gây tổn thương thần kinh và đau quá mẫn. J Biol Chem. 2007; 282: 14975 – 14983. [PubMed]
Kindt KS, Viswanath V, Macpherson L, Quast K, Hu H, Patapoutian A, Schafer WR. Caenorhabditis elegans TRPA-1 chức năng trong mechanosensation. Nat Neurosci. 2007; 10: 568 – 577. [PubMed]
Kirschstein T, Busselberg D, Treede RD. Coexpression của nhiệt-evoked và capsaicin-evoked inward currents trong sâu sắc disociated rễ chuột rễ hạch thần kinh. Neurosci Lett. 1997; 231: 33 – 36. [PubMed]
Knabl J, Witschi R, Hosl K, Reinold H, Zeilhofer UB, Ahmadi S, Brockhaus J, Sergejeva M, Hess A, Brune K, et al. Đảo ngược đau bệnh lý thông qua các phân nhóm thụ thể cột sống GABAA cụ thể. Thiên nhiên. 2008; 451: 330 – 334. [PubMed]
Kobayashi K, Yamanaka H, Fukuoka T, Đại Y, Obata K, Noguchi K. P2Y12 tăng cường thụ thể trong microglia được kích hoạt là một cửa ngõ của tín hiệu p38 và đau thần kinh. J Neurosci. 2008; 28: 2892 – 2902. [PubMed]
Kwan KY, Allchorne AJ, Vollrath MA, Christensen AP, Zhang DS, Woolf CJ, Corey DP. TRPA1 góp phần vào sự nociception lạnh, cơ học và hóa học nhưng không cần thiết cho sự truyền tải tế bào tóc. Neuron. 2006; 50: 277 – 289. [PubMed]
Kwan KY, Glazer JM, Corey DP, Rice FL, Stucky CL. TRPA1 điều chỉnh sự vận chuyển cơ học trong các tế bào thần kinh cảm giác da. J Neurosci. 2009; 29: 4808 – 4819. [PMC bài viết miễn phí] [PubMed]
Latremoliere A, Woolf CJ. Nhạy cảm trung tâm: một máy phát triển của quá mẫn cảm đau do dẻo thần kinh trung ương. J Đau. 2009; 10: 895 – 926. [PMC bài viết miễn phí] [PubMed]
Lawson JJ, McIlwrath SL, Woodbury CJ, Davis BM, Koerber HR. TRPV1 không giống như TRPV2 bị hạn chế đối với một nhóm nhỏ các nociceptors da nhạy cảm về mặt cơ học phản ứng với nhiệt. J Đau. 2008; 9: 298 – 308. [PMC bài viết miễn phí] [PubMed]
Leffler A, Linte RM, Nau C, Reeh P, Babes A. Một kênh kích hoạt nhiệt độ cao trong các tế bào thần kinh hạch rễ nuôi cấy lưng giống như TRPV2 và bị chặn bởi gadolinium. Eur J Neurosci. 2007; 26: 12 – 22. [PubMed]
Liedtke W, Choe Y, Marti-Renom MA, Bell AM, Denis CS, Sali A, Hudspeth AJ, Friedman JM, Heller S. Vanilloid liên quan đến thụ thể osmotically kích hoạt kênh (VR-OAC), một osmoreceptor vertebrate ứng cử viên. Ô. 2000; 103: 525 – 535. [PMC bài viết miễn phí] [PubMed]
Liedtke W, Friedman JM. Quy định thẩm thấu bất thường trong trpv4 - / - chuột. Proc Natl Acad Sci Mỹ A. 2003; 100: 13698 – 13703. [PMC bài viết miễn phí] [PubMed]
Lin YW, Cheng CM, Leduc PR, Chen CC. Hiểu sự vận chuyển cơ học thần kinh cảm giác thông qua kiểm soát ma trận đàn hồi cục bộ. PLoS One. 2009; 4: e4293. [PMC bài viết miễn phí] [PubMed]
Lindia JA, McGowan E, Jochnowitz N, Abbadie C. Cảm ứng biểu hiện CX3CL1 trong các tế bào hình sao và CX3CR1 trong microglia trong tủy sống của mô hình chuột đau thần kinh. J Đau. 2005; 6: 434 – 438. [PubMed]
Liu XJ, Gingrich JR, Vargas-Caballero M, Đồng YN, Sengar A, Beggs S, Wang SH, Ding HK, Frankland PW, Salter MW. Điều trị đau do viêm và đau thần kinh bằng cách tách Src khỏi phức hợp thụ thể NMDA. Nat Med. 2008; 14: 1325 – 1332. [PMC bài viết miễn phí] [PubMed]
Lumpkin EA, Caterina MJ. Cơ chế truyền tải giác quan trong da. Thiên nhiên. 2007; 445: 858 – 865. [PubMed]
Luo ZD, Chaplan SR, Higuera ES, Sorkin LS, Stauderman KA, Williams ME, Yaksh TL. Điều chỉnh tăng trưởng của hạch bạch huyết lưng (alpha) 2 (delta) tiểu đơn vị kênh canxi và mối tương quan của nó với allodynia trong chuột bị tổn thương thần kinh cột sống. J Neurosci. 2001; 21: 1868 – 1875. [PubMed]
Macpherson LJ, Dubin AE, Evans MJ, Marr F, Schultz PG, Cravatt BF, Patapoutian A. Các hợp chất độc hại kích hoạt các kênh ion TRPA1 thông qua sự thay đổi cộng hóa trị của cysteine. Thiên nhiên. 2007; 445: 541 – 545. [PubMed]
Madrid R, de la Pena E, Donovan-Rodriguez T, Belmonte C, Viana F. Biến số của các tế bào thần kinh cảm lạnh sinh nhiệt lạnh sinh học được xác định bằng sự cân bằng giữa các kênh kali TRPM8 và Kv1. J Neurosci. 2009; 29: 3120 – 3131. [PubMed]
Malan TP, Mata HP, Porreca F. Cột sống GABA (A) và GABA (B) dược lý thụ thể trong một mô hình chuột của đau thần kinh. Gây mê. 2002; 96: 1161 – 1167. [PubMed]
Malmberg AB, Chen C, Tonegawa S, Basbaum AI. Bảo tồn đau cấp tính và giảm đau thần kinh ở chuột thiếu PKCgamma. Khoa học. 1997; 278: 279 – 283. [PubMed]
Mandadi S, Sokabe T, Shibasaki K, Katanosaka K, Mizuno A, Moqrich A, Patapoutian A, Fukumi-Tominaga T, Mizumura K, Tominaga M. TRPV3 trong keratinocytes truyền thông tin nhiệt độ tới nơron cảm giác 2009 [PMC free article] [PubMed]
Materazzi S, Nassini R, Andre E, Campi B, Amadesi S, Trevisani M, Bunnett NW, Patacchini R, Geppetti P. Các chất chuyển hóa axit béo phụ thuộc Cox gây đau do kích hoạt thụ thể kích thích TRPA1. Proc Natl Acad Sci Mỹ A. 2008; 105: 12045 – 12050. [PMC bài viết miễn phí] [PubMed]
Matta JA, Cornett PM, Miyares RL, Abe K, Sahibzada N, Ahern GP. Thuốc gây mê tổng quát kích hoạt kênh ion nociceptive để tăng cường đau và viêm. Proc Natl Acad Sci Mỹ A. 2008; 105: 8784 – 8789. [PMC bài viết miễn phí] [PubMed]
McKemy DD, Neuhausser WM, Julius D. Xác định một thụ thể lạnh cho thấy một vai trò chung cho các kênh TRP trong quá trình nhiệt. Thiên nhiên. 2002; 416: 52 – 58. [PubMed]
McMahon SB, Bennett DLH, Bevan S. Các chất trung gian gây viêm và điều biến đau. Trong: McMahon SB, Koltzenburg M, biên tập viên. Sách giáo khoa về đau khổ của Wall và Melzack. Elsevier; 2008. pp. 49 – 72.
Melzack R, Tường PD. Cơ chế đau: một lý thuyết mới. Khoa học. 1965; 150: 971 – 979. [PubMed]
Messinger RB, Naik AK, Jagodic MM, Nelson MT, Lee WY, Choe WJ, Orestes P, Latham JR, Todorovic SM, Jevtovic-Todorovic V. Trong im lặng cơ thể của Ca (V) 3.2 T-loại kênh canxi trong tế bào thần kinh cảm giác làm giảm bớt chứng tăng sản ở chuột với bệnh lý thần kinh do tiểu đường do streptozocin gây ra. Đau đớn. 2009; 145: 184 – 195. [PMC bài viết miễn phí] [PubMed]
Milligan ED, Sloane EM, Watkins LR. Glia trong đau bệnh lý: một vai trò cho fractalkine. J Neuroimmunol. 2008; 198: 113 – 120. [PMC bài viết miễn phí] [PubMed]
Milligan ED, Zapata V, Chacur M, Schoeniger D, Biedenkapp J, O'Connor KA, Verge GM, Chapman G, Green P, Foster AC, et al. Bằng chứng cho thấy ribalkine ngoại sinh và nội sinh có thể gây kích thích nociceptive cột sống ở chuột. Eur J Neurosci. 2004; 20: 2294 – 2302. [PubMed]
Miraucourt LS, D song R, Voisin DL. Glycine ức chế rối loạn chức năng biến cảm ứng đau đớn thông qua PKCgamma interneurons. PLoS One. 2007; 2: e1116. [PMC bài viết miễn phí] [PubMed]
Mochizuki T, Sokabe T, Araki I, Fujishita K, Shibasaki K, Uchida K, Naruse K, Koizumi S, Takeda M, Tominaga M. Kênh cation TRPV4 làm trung gian phát triển Ca2 + dòng và phát hành ATP trong các tế bào tế bào nội mô chính. J Biol Chem. 2009; 284: 21257 – 21264. [PMC bài viết miễn phí] [PubMed]
Moore KA, Kohno T, Karchewski LA, Scholz J, Baba H, Woolf CJ. Chấn thương thần kinh ngoại vi một phần thúc đẩy một sự mất mát có chọn lọc ức chế GABAergic ở sừng lưng bề ngoài của tủy sống. J Neurosci. 2002; 22: 6724 – 6731. [PubMed]
Mueller KL, Hoon MA, Erlenbach I, Chandrashekar J, Zuker CS, Ryba NJ. Các thụ thể và logic mã hóa cho vị đắng. Thiên nhiên. 2005; 434: 225 – 229. [PubMed]
Muraki K, Iwata Y, Katanosaka Y, Ito T, Ohya S, Shigekawa M, Imaizumi Y. TRPV2 là một thành phần của các kênh cation nhạy cảm thẩm thấu trong tế bào cơ tim chuột. Circ Res. 2003; 93: 829 – 838. [PubMed]
Nagata K, Duggan A, Kumar G, Garcia-Anoveros J. Nociceptor và các tính chất đầu dò tế bào tóc của TRPA1, một kênh cho đau và thính giác. J Neurosci. 2005; 25: 4052 – 4061. [PubMed]
Nagy I, Rang H. Noxious nhiệt kích hoạt tất cả capsaicin nhạy cảm và cũng là một dân số phụ của các tế bào thần kinh hạch rễ không nhạy cảm capsaicin-insensitive. Khoa học thần kinh. 1999; 88: 995 – 997. [PubMed]
Nassar MA, Levato A, Stirling LC, Gỗ JN. Đau thần kinh phát triển bình thường ở chuột thiếu cả Na (v) 1.7 và Na (v) 1.8. Đau Mol. 2005; 1: 24. [PMC bài viết miễn phí] [PubMed]
Nassar MA, Stirling LC, Forlani G, Baker MD, Matthews EA, Dickenson AH, Gỗ JN. Nociceptor-cụ thể gen xóa cho thấy một vai trò quan trọng đối với Nav1.7 (PN1) trong đau cấp tính và viêm. Proc Natl Acad Sci Mỹ A. 2004; 101: 12706 – 12711. [PMC bài viết miễn phí] [PubMed]
Neumann S, Braz JM, Skinner K, Llewellyn-Smith IJ, Basbaum AI. Vô hại, không độc hại, đầu vào kích hoạt PKCgamma interneurons của sừng lưng cột sống thông qua sợi affelent myelinated. J Neurosci. 2008; 28: 7936 – 7944. [PMC bài viết miễn phí] [PubMed]
Nichols ML, Allen BJ, Rogers SD, Ghilardi JR, Honore P, Luger NM, Finke MP, Li J, Lappi DA, Simone DA, et al. Sự truyền nhiễm của nociception mãn tính bởi các tế bào thần kinh cột sống biểu hiện chất thụ thể P. Khoa học. 1999; 286: 1558 – 1561. [PubMed]
Noel J, Zimmermann K, Busserolles J, Deval E, Alloui A, Diochot S, Guy N, Borsotto M, Reeh P, Eschalier A, et al. Các kênh K + được kích hoạt bằng mechano TRAAK và TREK-1 kiểm soát cả nhận thức ấm và lạnh. EMBO J. 2009; 28: 1308 – 1318. [PMC bài viết miễn phí] [PubMed]
Obata K, Katsura H, Miyoshi K, Kondo T, Yamanaka H, Kobayashi K, Đại Y, Fukuoka T, Akira S, Noguchi K. Toll-like receptor 3 góp phần kích hoạt cột sống và xúc giác allodynia sau chấn thương thần kinh. J Neurochem 2008 [PubMed]
Olausson H, Cole J, Rylander K, McGlone F, Lamarre Y, Wallin BG, Kramer H, Wessberg J, Elam M, Bushnell MC, et al. Vai trò chức năng của các nhân viên xúc giác không khớp trong da lông người: phản ứng giao cảm và nội địa hóa cảm nhận. Exp Brain Res. 2008; 184: 135 – 140. [PubMed]
Trang AJ, Brierley SM, Martin CM, Martinez-Salgado C, Wemmie JA, Brennan TJ, Symonds E, Omari T, Lewin GR, Welsh MJ, et al. Kênh ion ASIC1 góp phần vào chức năng cơ khí nội tạng nhưng không phải là da. Gastroenterology. 2004; 127: 1739 – 1747. [PubMed]
Peier AM, Moqrich A, Hergarden AC, Reeve AJ, Andersson DA, Câu chuyện GM, Earley TJ, Dragoni I, McIntyre P, Bevan S, et al. Một kênh TRP cảm nhận kích thích lạnh và tinh dầu bạc hà. Ô. 2002a; 108: 705 – 715. [PubMed]
Peier AM, Reeve AJ, Andersson DA, Moqrich A, Earley TJ, Hergarden AC, Câu chuyện GM, Colley S, Hogenesch JB, McIntyre P, et al. Một kênh TRP nhạy cảm nhiệt biểu hiện trong các tế bào sừng. Khoa học. 2002b; 296: 2046 – 2049. [PubMed]
Perl ER. Ý tưởng về nỗi đau, một cái nhìn lịch sử. Nat Rev Neurosci. 2007; 8: 71 – 80. [PubMed]
Petrus M, Peier AM, Bandell M, Hwang SW, Huỳnh T, Olney N, Jegla T, Patapoutian A. Một vai trò của TRPA1 trong hyperalgesia cơ được tiết lộ bởi ức chế dược lý. Đau Mol. 2007; 3: 40. [PMC bài viết miễn phí] [PubMed]
Polgar E, Hughes DI, Arham AZ, Todd AJ. Mất tế bào thần kinh từ laminas I-III của sừng lưng cột sống là không cần thiết cho sự phát triển của allodynia xúc giác trong mô hình tổn thương thần kinh bị tổn thương của đau thần kinh. J Neurosci. 2005; 25: 6658 – 6666. [PubMed]
Porreca F, Ossipov MH, Gebhart GF. Đau mãn tính và giảm dần thuận lợi giảm dần. Xu hướng Neurosci. 2002; 25: 319 – 325. [PubMed]
Giá MP, Lewin GR, McIlwrath SL, Cheng C, Xie J, Heppenstall PA, Stucky CL, Mannsfeldt AG, Brennan TJ, Drummond HA, et al. Kênh natri của động vật có vú BNC1 là cần thiết cho cảm giác bình thường. Thiên nhiên. 2000; 407: 1007 – 1011. [PubMed]
Giá MP, McIlwrath SL, Xie J, Cheng C, Qiao J, Tarr DE, Sluka KA, Brennan TJ, Lewin GR, Welsh MJ. Kênh cation DRASIC góp phần vào việc phát hiện cảm ứng da và kích thích acid ở chuột. Neuron. 2001; 32: 1071 – 1083. [PubMed]
Rau KK, Giang N, Johnson RD, Cooper BY. Nhạy cảm nhiệt trong da và cơ bắp nociceptors thể hiện sự kết hợp riêng biệt của TRPV1 và TRPV2 protein. J Neurophysiol. 2007; 97: 2651 – 2662. [PubMed]
Rauck RL, Wallace MS, Burton AW, Kapural L, Bắc JM. Ziconotide trong vỏ cho đau thần kinh: một đánh giá. Đau thực hành. 2009; 9: 327 – 337. [PubMed]
Reid G, Flonta ML. Sinh lý học. Dòng điện lạnh trong các nơron chịu nhiệt. Thiên nhiên. 2001; 413: 480. [PubMed]
Ren K, Dubner R. Neuron-glia nhiễu xuyên âm nghiêm trọng: vai trò trong quá mẫn cảm đau. Curr Opin Anaesthesiol. 2008; 21: 570 – 579. [PMC bài viết miễn phí] [PubMed]
Ritner HL, Machelska H, Stein C. Hệ thống miễn dịch Đau và giảm đau. Trong: Basbaum AI, Bushnell M, biên tập viên. Khoa học của đau. 2009. pp. 407 – 427.
Roza C, Puel JL, Kress M, Baron A, Diochot S, Lazdunski M, Waldmann R. Knockout của kênh ASIC2 ở chuột không làm giảm quá trình cơ khí hóa da, cơ chế nội tạng và thính giác nội tạng. J Physiol. 2004; 558: 659 – 669. [PMC bài viết miễn phí] [PubMed]
Scherrer G, Imamachi N, Cao YQ, Contet C, Mennicken F, O'Donnell D, Kieffer BL, Basbaum AI. Phân tách các cơ chế thụ thể opioid kiểm soát đau cơ và nhiệt. Ô. 2009; 137: 1148 – 1159. [PMC bài viết miễn phí] [PubMed]
Schmidt R, Schmelz M, Forster C, Ringkamp M, Torebjork E, Handwerker H. Novel các lớp C nhạy cảm và không phản ứng với các nociceptors trong da người. J Neurosci. 1995; 15: 333 – 341. [PubMed]
Sivilotti L, Woolf CJ. Sự đóng góp của các thụ thể GABAA và glycine đến sự nhạy cảm trung tâm: sự mất cân bằng và kích thích tất cả chứng mất ngủ trong tủy sống. J Neurophysiol. 1994; 72: 169 – 179. [PubMed]
Snider WD, McMahon SB. Giải quyết nỗi đau ở nguồn: những ý tưởng mới về người nuôi nociceptors. Neuron. 1998; 20: 629 – 632. [PubMed]
Câu chuyện GM, Peier AM, Reeve AJ, Eid SR, Mosbacher J, Hricik TR, Earley TJ, Hergarden AC, Andersson DA, Hwang SW, et al. ANKTM1, một kênh giống như TRP thể hiện trong các tế bào thần kinh nociceptive, được kích hoạt bởi nhiệt độ lạnh. Ô. 2003; 112: 819 – 829. [PubMed]
Strotmann R, Harteneck C, Nunnenmacher K, Schultz G, Nhà máy TD. OTRPC4, một kênh cation không chọn lọc tạo ra độ nhạy với độ thẩm thấu ngoại bào. Nat Cell Biol. 2000; 2: 695 – 702. [PubMed]
Sugai E, Morimitsu Y, Iwasaki Y, Morita A, Watanabe T, Kubota K. Pungent phẩm chất của các hợp chất liên quan đến sanshool được đánh giá bằng một thử nghiệm cảm giác và kích hoạt chuột TRPV1. Biosci Biotechnol Biochem. 2005; 69: 1951 – 1957. [PubMed]
Suzuki M, Watanabe Y, Oyama Y, Mizuno A, Kusano E, Hirao A, Ookawara S. Bản địa hóa kênh cơ học TRPV4 trong da chuột. Neurosci Lett. 2003; 353: 189 – 192. [PubMed]
Swayne LA, Bourinet E. Các kênh canxi có điện áp trong đau mãn tính: vai trò mới nổi của mối nối thay thế. Pflugers Arch. 2008; 456: 459 – 466. [PubMed]
Takahashi A, Gotoh H. Các dòng tế bào toàn cơ học trong tế bào thần kinh chuột somatosensory nuôi cấy. Brain Res. 2000; 869: 225 – 230. [PubMed]
Tanga FY, Nutile-McMenemy N, DeLeo JA. Vai trò CNS của thụ thể giống như Toll 4 trong thần kinh bẩm sinh và đau thần kinh đau đớn. Proc Natl Acad Sci Mỹ A. 2005; 102: 5856 – 5861. [PMC bài viết miễn phí] [PubMed]
Tominaga M, Caterina MJ, Malmberg AB, Rosen TA, Gilbert H, Skinner K, Raumann BE, Basbaum AI, Julius D. Các thụ thể capsaicin nhân bản vô tính tích hợp nhiều kích thích gây đau. Neuron. 1998; 21: 531 – 543. [PubMed]
Torsney C, MacDermott AB. Disinhibition mở ra cánh cửa để bệnh lý đau báo hiệu trong neurotin neurokinin tế bào thần kinh thể hiện thụ thể 1 trong tủy sống chuột. J Neurosci. 2006; 26: 1833 – 1843. [PubMed]
Tozaki-Saitoh H, Tsuda M, Miyata H, Ueda K, Kohsaka S, Inoue K. P2Y12 thụ thể trong microglia cột sống là cần thiết cho đau thần kinh sau chấn thương dây thần kinh ngoại vi. J Neurosci. 2008; 28: 4949 – 4956. [PubMed]
Trevisani M, Siemens J, Materazzi S, Bautista DM, Nassini R, Campi B, Imamachi N, Andre E, Patacchini R, Cottrell GS, et al. 4-Hydroxynonenal, một aldehyde nội sinh, gây đau và viêm thần kinh do kích hoạt thụ thể gây kích ứng TRPA1. Proc Natl Acad Sci Mỹ A. 2007; 104: 13519 – 13524. [PMC bài viết miễn phí] [PubMed]
Tsuda M, Shigemoto-Mogami Y, Koizumi S, Mizokoshi A, Kohsaka S, Salter MW, Inoue K. P2X4 thụ thể gây ra trong tất cả các cơn đau dây thần kinh cột sống sau chấn thương thần kinh. Thiên nhiên. 2003; 424: 778 – 783. [PubMed]
Ulmann L, Hatcher JP, Hughes JP, Chaumont S, Green PJ, Conquet F, Buell GN, Reeve AJ, Chessell IP, Rassendren F. Điều chỉnh các thụ thể P2X4 trong microglia cột sống sau chấn thương dây thần kinh ngoại vi làm trung gian giải phóng BDNF và đau thần kinh. J Neurosci. 2008; 28: 11263 – 11268. [PubMed]
Xác minh GM, Milligan ED, Maier SF, Watkins LR, Naeve GS, Foster AC. Fractalkine (CX3CL1) và thụ thể fractalkine (CX3CR1) phân bố trong tủy sống và hạch rễ lưng dưới điều kiện đau cơ bản và đau thần kinh. Eur J Neurosci. 2004; 20: 1150 – 1160. [PubMed]
Viana F, de la Pena E, Belmonte C. Độ đặc hiệu của quá trình truyền nhiệt lạnh được xác định bằng biểu hiện kênh ion khác biệt. Nat Neurosci. 2002; 5: 254 – 260. [PubMed]
Wetzel C, Hu J, Riethmacher D, Benckendorff A, Harder L, Eilers A, Moshourab R, Kozlenkov A, Labuz D, Caspani O, et al. Một protein miền stomatin cần thiết cho cảm giác cảm ứng trên chuột. Thiên nhiên. 2007; 445: 206 – 209. [PubMed]
Woodbury CJ, Zwick M, Wang S, Lawson JJ, Caterina MJ, Koltzenburg M, Albers KM, Nhân viên Koerber, Davis BM. Nociceptors thiếu TRPV1 và TRPV2 có phản ứng nhiệt bình thường. J Neurosci. 2004; 24: 6410 – 6415. [PubMed]
Woolf CJ. Bằng chứng cho một thành phần trung tâm của quá mẫn sau chấn thương đau. Thiên nhiên. 1983; 306: 686 – 688. [PubMed]
Yagi J, Wenk HN, Naves LA, McCleskey EW. Dòng chảy bền vững thông qua các kênh ion ASIC3 ở những thay đổi pH khiêm tốn xảy ra trong thời gian thiếu máu cục bộ cơ tim. Circ Res. 2006; 99: 501 – 509. [PubMed]
Yaksh TL. Tương quan hành vi và tự trị của xúc giác gợi lên allodynia do ức chế glycine cột sống: tác động của hệ thống thụ thể điều hòa và chất đối kháng acid amin kích thích. Đau đớn. 1989; 37: 111 – 123. [PubMed]
Yang Y, Wang Y, Li S, Xu Z, Li H, Ma L, Fan J, Bu D, Liu B, Fan Z, et al. Các đột biến trong SCN9A, mã hóa một tiểu đơn vị alpha kênh alpha, ở những bệnh nhân bị đau cơ ban đầu. J Med Genet. 2004; 41: 171 – 174. [PMC bài viết miễn phí] [PubMed]
Zamponi GW, Lewis RJ, Todorovic SM, Arneric SP, Snutch TP. Vai trò của các kênh canxi điện áp-gated trong con đường đau tăng dần. Brain Res Rev. 2009; 60: 84 – 89. [PMC bài viết miễn phí] [PubMed]
Zhuang ZY, Kawasaki Y, Tân PH, Wen YR, Huang J, Ji RR. Vai trò của con đường MAPX CX3CR1 / p38 trong vi mô cột sống cho sự phát triển của đau thần kinh sau sự phân tách do tổn thương dây thần kinh của gãy xương. Brain Behav Immun. 2007; 21: 642 – 651. [PMC bài viết miễn phí] [PubMed]
Zimmermann K, Leffler A, Babes A, Cendan CM, Carr RW, Kobayashi J, Nau C, Gỗ JN, Reeh PW. Kênh natri thần kinh cảm giác Nav1.8 là điều cần thiết cho cơn đau ở nhiệt độ thấp. Thiên nhiên. 2007; 447: 855 – 858. [PubMed]
Zurborg S, Yurgionas B, Jira JA, Caspani O, Heppenstall PA. Kích hoạt trực tiếp kênh ion TRPA1 bằng Ca2 + Nat Neurosci. 2007; 10: 277 – 279. [PubMed]

Các chủ đề khác: Đau lưng
đau lưng là một trong những nguyên nhân phổ biến nhất cho tình trạng khuyết tật và những ngày bỏ lỡ tại nơi làm việc trên toàn thế giới. Như một vấn đề của thực tế, đau lưng đã được coi là lý do phổ biến thứ hai cho các chuyến thăm văn phòng bác sĩ, đông hơn chỉ bởi nhiễm trùng đường hô hấp trên. Khoảng 80 phần trăm dân số sẽ trải qua một số loại đau lưng ít nhất một lần trong suốt cuộc đời của họ. Cột sống là một cấu trúc phức tạp gồm xương, khớp, dây chằng và cơ bắp, cùng với các mô mềm khác. Bởi vì điều này, chấn thương và / hoặc tình trạng tăng nặng, chẳng hạn như đĩa đệm thoát vị, cuối cùng có thể dẫn đến các triệu chứng đau lưng. Thương tích thể thao hoặc chấn thương tai nạn ô tô thường là nguyên nhân thường gặp nhất gây đau lưng, tuy nhiên, đôi khi cử động đơn giản nhất có thể có kết quả đau đớn. May mắn thay, lựa chọn điều trị thay thế, chẳng hạn như chăm sóc chỉnh hình, có thể giúp giảm đau lưng thông qua việc sử dụng điều chỉnh cột sống và thao tác thủ công, cuối cùng là cải thiện giảm đau.

EXTRA QUAN TRỌNG CHỦ ĐỀ: Quản lý đau lưng thấp
CÁC CHỦ ĐỀ KHÁC: EXTRA EXTRA: Điều trị & Đau mãn tính
Đăng tuyên bố từ chối trách nhiệm
Phạm vi hành nghề chuyên nghiệp *
Thông tin ở đây về "Cơ chế đau cấp tính và đau mãn tính" không nhằm mục đích thay thế mối quan hệ trực tiếp với chuyên gia chăm sóc sức khỏe đủ điều kiện hoặc bác sĩ được cấp phép và không phải là lời khuyên y tế. Chúng tôi khuyến khích bạn đưa ra quyết định chăm sóc sức khỏe dựa trên nghiên cứu và quan hệ đối tác của bạn với chuyên gia chăm sóc sức khỏe đủ điều kiện.
Thông tin Blog & Thảo luận Phạm vi
Phạm vi thông tin của chúng tôi được giới hạn ở Chiropractic, cơ xương khớp, châm cứu, thuốc vật lý, sức khỏe, góp phần căn nguyên rối loạn nội tạng trong các bài thuyết trình lâm sàng, động lực học phản xạ somatovisceral liên quan đến lâm sàng, các phức hợp hạ lưu, các vấn đề sức khỏe nhạy cảm và / hoặc các bài báo, chủ đề và thảo luận về y học chức năng.
Chúng tôi cung cấp và trình bày cộng tác lâm sàng với các chuyên gia từ nhiều lĩnh vực khác nhau. Mỗi chuyên gia được điều chỉnh bởi phạm vi hành nghề chuyên nghiệp và thẩm quyền cấp phép của họ. Chúng tôi sử dụng các phác đồ chăm sóc sức khỏe & sức khỏe chức năng để điều trị và hỗ trợ chăm sóc các chấn thương hoặc rối loạn của hệ thống cơ xương.
Các video, bài đăng, chủ đề, chủ đề và thông tin chi tiết của chúng tôi đề cập đến các vấn đề, vấn đề và chủ đề lâm sàng liên quan và trực tiếp hoặc gián tiếp hỗ trợ phạm vi hành nghề lâm sàng của chúng tôi.*
Văn phòng của chúng tôi đã cố gắng một cách hợp lý để cung cấp các trích dẫn hỗ trợ và đã xác định các nghiên cứu có liên quan hỗ trợ các bài đăng của chúng tôi. Chúng tôi cung cấp các bản sao của các nghiên cứu hỗ trợ có sẵn cho các hội đồng quản lý và công chúng theo yêu cầu.
Chúng tôi hiểu rằng chúng tôi bao gồm các vấn đề cần giải thích thêm về cách nó có thể hỗ trợ trong một kế hoạch chăm sóc hoặc phác đồ điều trị cụ thể; do đó, để thảo luận thêm về vấn đề trên, vui lòng hỏi Tiến sĩ Alex Jimenez, DC, hoặc liên hệ với chúng tôi tại 915-850-0900.
Chúng tôi ở đây để giúp bạn và gia đình bạn.
Blessings
Tiến sĩ Alex Jimenez A.D, MSACP, RN*, CCST, IFMCP*, CIFM*, ATN*
email: coach@elpasofeftalmedicine.com
Được cấp phép là Bác sĩ Chiropractic (DC) tại Texas & New Mexico*
Giấy phép Texas DC # TX5807, Giấy phép New Mexico DC # NM-DC2182
Được cấp phép làm Y tá đã Đăng ký (RN*) tại Florida
Giấy phép Florida Giấy phép RN # RN9617241 (Số kiểm soát 3558029)
Trạng thái nhỏ gọn: Giấy phép đa bang: Được phép hành nghề tại Hoa Kỳ*
Hiện đã trúng tuyển: ICHS: MSN* FNP (Chương trình bác sĩ y tá gia đình)
Tiến sĩ Alex Jimenez DC, MSACP, RN* CIFM*, IFMCP*, ATN*, CCST
Danh thiếp kỹ thuật số của tôi
 Một lần nữa, chúng tôi chào đón bạn¸
Một lần nữa, chúng tôi chào đón bạn¸
